"Основы физиологии высшей нервной деятельности" - читать интересную книгу автора (Коган Александр Борисович)
Глава 3 ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ПАМЯТИ
Эффективность приспособительного поведения животных и успех разумной деятельности человека в значительной мере зависят от того, в какой мере животные используют свой жизненный опыт, а человек — полученные ранее знания для того, чтобы принимать правильные решения и избегать ошибочных. Эти знания извлекаются из
Различия механизмов осуществления этих проявлений памяти наглядно демонстрируются врачебными наблюдениями за больными, у которых разные поражения мозга вызывали различные нарушения памяти. Например, описаны случаи, когда человек запоминает с трудом, но помнит долго или может запомнить лишь на очень короткое время или с большим трудом вспоминает то, что он легко запомнил.
Таким образом,
Давно замечено, что при некоторых заболеваниях мозга теряется способность запоминать события, происходящие во время болезни, хотя все бывшее ранее больной хорошо помнит. Эти явления в резкой форме были вначале обнаружены и описаны при тяжелой форме алкоголизма (С.С. Корсаков, 1880). Больной не был в состоянии запомнить происходящие события, но мог помнить, что происходило с ним раньше. Такую потерю памяти на недавние события наблюдали также у людей, переживших сотрясение мозга, при его очаговых поражениях, особенно в области височных долей, затрагивающих гиппокамп. Психологи отмечали сходные явления забывания событий, предшествовавших нервному потрясению. При этом память на все более давние события полностью сохранялась.
Такие явления
Нарушения краткосрочной памяти типа ретроградной амнезии оказалось возможным воспроизвести в экспериментах на животных. Для этого достаточно было сразу после обучения подвергнуть мозг сильному воздействию и только что приобретенное знание исчезало, хотя ранее приобретенные навыки сохранялись. В экспериментах такими воздействиями были различные фармакологические средства, наркотики, переохлаждение мозга, нарушение его кислородного снабжения и другие средства. Однако чаще всего применяли электрическое раздражение мозга, вызывающее судорожные припадки.
На рис. 20 приведены результаты опыта, в котором у мышей вырабатывали условный оборонительный рефлекс избегания выхода на площадку, находящуюся под напряжением.
Получив удар током, мышь избегала выходить на эту площадку. Учитывали поведение мыши в первые 30 с после выхода на площадку. Как видно из рис. 20, если эфирный наркоз был дан сразу или на протяжении не более 10 мин после удара током на площадке, то условный рефлекс ее избегания был нарушен, мышь продолжала выбегать на площадку. Если наркоз был дан позже, спустя 16–20 мин, то он разрушал рефлекс не полностью, а наркотизация через 24 мин вообще не влияла на него.
Многие эксперименты показывали возможность избирательно выключать кратковременную память, не нарушая долговременную, и избирательно воздействовать на долгосрочную память, не нарушая краткосрочную. Такое действие оказывают, например, некоторые антибиотики, угнетающие синтез белка. Из этих данных был сделан вывод, что краткосрочная и долгосрочная память имеют разную природу и регулируются разными механизмами. Однако многие воздействия, выключающие краткосрочную память, как, например, нанесение на поверхность мозга раствора хлористого калия, нарушали и долговременную память. Вместе с тем описаны случаи, когда ретроградная амнезия через некоторое время проходила сама или снималась действием тока более слабого, чем применяемый в качестве подкрепления при выработке исследуемого рефлекса.
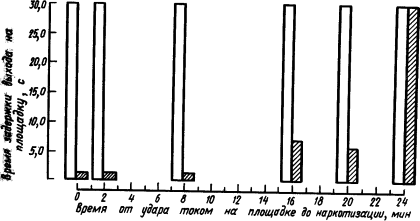 |
Рис. 20. Влияние наркоза на закрепление у мышей рефлексов избегания выхода на площадку, находящуюся под напряжением (по И. Абт и др.):
белыми столбиками изображено проявление рефлекса до наркотизации, заштрихованными — после наркотизации в разные сроки; остальные объяснения см. в тексте
На основании опытов с обезьянами было высказано предположение, что в условно-рефлекторной деятельности проявляются оба вида памяти: при отсроченных условных рефлексах — преимущественно краткосрочная, при следовых условных рефлексах — долгосрочная. Поэтому
Возможные механизмы краткосрочной памяти. Наиболее распространено предположение, что краткосрочная память реализуется в виде многократной циркуляции потоков импульсов по замкнутым цепям нервных клеток. Основанием для такой гипотезы
Однако дальнейшие исследования показали, что в круговых путях нервных сетей находятся не только возбуждающие, но и тормозящие клетки, которые препятствуют «зацикливанию» циркуляции импульсов. На рис. 21 показана такая схема сети, в которой клетка
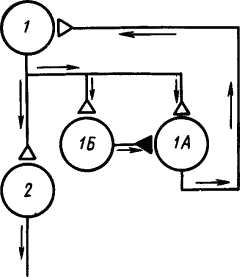 |
Рис. 21. Нервная цепь с замкнутыми нейронными цепями, по которым могут циркулировать потоки импульсов (по Э. Кенделу):
Краткосрочная память не требует для своей реализации синтеза информационных (матричных) молекул РНК и ДНК. Она и не могла бы обеспечиваться этими биохимическими процессами как генетическая память, поскольку включение в полимер ДНК или РНК нуклеотидов происходит в сотни и тысячи раз медленнее, чем это необходимо для запечатления в них полной информации о текущих событиях. Вместе с тем при любом понимании ее природы, в том числе и реверберационном, речь идет о протекании потоков импульсов по новым для них путям, т.е. о временной
Для объяснения механизма синаптической проводимости была предложена гипотеза о взаимодействии медиатора ацетилхолина с рецепторными белками постсинаптической мембраны и Na+, K+-АТФ-азой при разных концентрациях ионов Na и Ca. При интенсивной импульсации выброшенный из везикул пресинаптической мембраны ацетилхолин связывается с рецепторными белками постсинаптической мембраны, подавляет деятельность ионных насосов, деполяризует мембрану и выключает Na+, K+-АТФ-азу. Однако возникающие при этом потенциалы возбуждения и уменьшение отношения Na+ и K+ приводят к высвобождению ацетилхолина и последующему разрушению его холинэстеразой, что может восстанавливать активность Na+, К+-АТФ-азы на более высоком уровне.
Временное повышение синаптической проводимости связывают также с высвобождением Ca2+ под действием нервных импульсов в пресинаптических окончаниях, который вызывает усиленное сокращение везикул и выбрасывание медиатора, или с повышенной чувствительностью белков деполяризованной мембраны к протеолитическим ферментам, делающим ее более проходимой для медиатора. В пользу последнего предположения свидетельствует возрастание активности протеаз в ткани мозга животных при выработке условного рефлекса.
Исходя из результатов электрофизиологических исследований было высказано мнение, что краткосрочная память образуется за счет
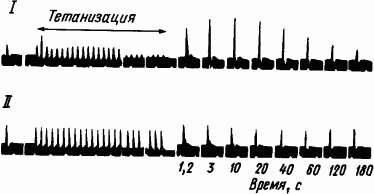 |
Рис. 22. Посттетаническое временно́е облегчение проведения через звездчатый ганглий кошки (по М. Ларраби, Д. Бронк):
Несмотря на различия предположений о природе краткосрочной памяти, их объединяет рассмотрение механизма памяти как
В отличие от краткосрочной долгосрочная память хранит следы пережитых событий неопределенно долго. Собственно она и является носителем жизненного опыта животного; знаний, приобретенных человеком. О субстрате и способе закрепления долгосрочных памятных следов существуют разные мнения.
Долгосрочная память как возникновение новых межклеточных связей и улучшение проведения по уже существующим связям. Не вызывает сомнений, что в основе всякого обучения лежит процесс проторения путей возбуждения. Наглядной иллюстрацией того, что в результате обучения потоки нервных импульсов легче и быстрее преодолевают синаптическую задержку, может служить модельный опыт с тараканом: после ампутации передних лапок, которыми он чистит усики, таракан обучается пользоваться для этой цели задними лапками. При этом время синаптической задержки моторных импульсов к задним лапкам резко уменьшается (рис. 23).
Исследование динамики активности нейронов виноградной улитки при выработке оборонительного условного рефлекса показало, что повышение эффективности синапсов происходит на входах командных нейронов, реализующих защитные движения.
При выработке условного рефлекса постановки лапы кошки на подставку исследовали реакции ЭЭГ и нейронной активности в моторной коре. Наблюдаемое при этом их единообразие в случае применения различных по характеру и интенсивности условных раздражителей привело к заключению, что сигнал лишь запускает программу ответного двигательного акта. Это происходит, когда наступает повышение эффективности синапсов на входах нейронов — генераторов команд.
 |
Рис. 23. Укорочение времени прохождения через метаторакальный ганглий таракана нервных импульсов, приводящих в движение его задние лапки, которыми он научился чистить усики после ампутации передних лапок (по Д. Люко, А. Аранда):
Давно высказанное предположение, что памятный след закрепляется «прорастанием» дополнительных нервных окончаний (Р. Кахал, 1911), позже получило новые подтверждения. Оказалось, что так называемые шипики, несущие синапсы на дендритах корковых клеток, появляются именно в возрасте, когда развивается условно-рефлекторная деятельность. Дополнительные указания на роль шипиков в мехаизмзах памяти дали эксперименты, показавшие интенсивное развитие их синаптического аппарата у животных в результате выработки условных рефлексов. К такому пониманию природы долгосрочной памяти приводит и концепция участия генетического аппарата в пластическом обеспечении функций нервных клеток, согласно которой их интенсивная деятельность при обучении вызывает усиление синтеза белка и процессов роста, в частности, образующих новые синаптические связи. Наконец, зависимость количества синапсов (по показателям электронной микроскопии) от интенсивности функционирования нервных механизмов продемонстрирована в экспериментах с повышением активности синапсов путем предъявления сложных задач или ее снижения путем сенсорной депривации.
По другим предположениям долгосрочная память реализуется
Эффективность передачи возбуждения в синапсах в значительной мере зависит от их функционального состояния, определяемого уровнем деятельности. Показано, что
Одним из механизмов повышения эффективности синапсов могут быть процессы, проявляющиеся в так называемой
Таблица 3. Возможные механизмы устойчивого повышения проводимости в синапсах (по С. Барондес в модификации И.П. Ашмарина)
1. Область изменений в синапсе
2. Состояние синаптической щели
3. Характер изменений
4. Молекулярный механизм
1. Пресинаптическая мембрана
2. Сужение
3. Разрастание нервных окончаний
4. Усиленный синтез белков при запоминании, в дальнейшем их обновление
1. —
2. Без изменения
3. Усиление синтеза и выхода медиатора
4. Дерегрессия части генов медиатора, повышение активности систем трансляции и РНК медиатора
Усиление синтеза медиатора и облегчение выхода из везикул
1. Постсинаптическая мембрана
2. Без изменений
3. Увеличение числа и чувствительности рецепторов
4. Усиленный синтез белков и липидов при запоминании, в дальнейшем их обновление
1. —
2. —
3. Повышение проницаемости и уменьшение скорости расщепления медиатора
4. Измерение соотношений скоростей синтеза компонентов или образование новых компонентов рецептора при запоминании, в дальнейшем поддержание новых отношений. Репрессия генов энзима, расщепляющего медиатор, дерепрессия генов ингибитора этого энзима
1. Локализация не установлена
2. Не известно
3. Повышение проводимости синапса за счет включения неизвестного вещества
4. Усиленный синтез веществ типа антител или других факторов межклеточного «узнавания» (например, полипептиды — коннекторы), при запоминании, в дальнейшем их обновление
Усиленный синтез некоторых антигенов компонента синапса, вызывающих образование соответствующих антител при запоминании, в дальнейшем их обновление
Анализ этих явлений нейронной активности при выработке локальной условной реакции вздрагивания приводит к выводу о том, что для синаптического проторения путей временной связи необходима кроме конвергенции сигнального и безусловного стимулов активация системы, вызывающей модификацию синапсов, что зависит от биологической значимости образования условной реакции. На изолированном нейроне виноградной улитки получены условные изменения свойств его электровозбудимой мембраны путем сочетания стимулов, приложенных к двум ее локусам, в результате чего между ними возникала временная связь.
При изучении возможных причин стойкого повышения синаптической передачи было обращено внимание на
Сопоставление данных о взаимосвязанной динамике ацетилхолина и холинэстеразы привело к заключению, что повышенная проводимость может возникать и устойчиво поддерживаться некоторым равновесным отношением их синтеза. Избыточная активность холинэстеразы без возрастания количества ацетилхолина не позволяет ему достичь концентрации, необходимой для деполяризации постсинаптической мембраны синапса, а увеличение количества ацетилхолина при малой активности холинэстеразы приводит к глубокой неколеблющейся деполяризации — в обоих случаях проведение импульсов через синапс оказывается блокированным. Существует предположение, что ацетилхолин может транспортироваться через синаптическую щель по транссинаптическим тяжам, которые образуются из соединения белковых мицелл, формирующихся на поверхности пре- и постсинаптической мембран.
Кроме холинэргических имеются и иные синапсы, где медиаторами служат другие вещества. С учетом всех этих сведений была составлена обобщенная схема стойкого облегчения медиаторных процессов в синапсе. На рис. 24,
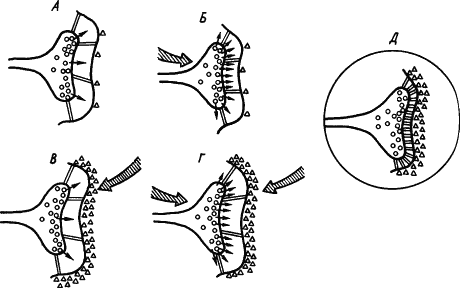 |
Рис. 24. Развитие процессов стойкого облегчения проводимости сигнала (объяснение см. в тексте) (по В.В. Дергачеву):
кружками обозначены везикулы медиатора, треугольниками — фермент, расцепляющий медиатор
Долгосрочная память как стойкое изменение химизма нервных клеток. Проведение сигналов от клетки к клетке, вызывающее активацию нейронных популяций, происходит в синапсах, однако функциональное состояние синапсов и свойства их деятельности в значительной мере определяются процессами, происходящими в соме клетки, в ее нейроплазме и ядре. Общеизвестна роль сомы в трофическом обеспечении передачи импульсов по путям их проведения и зависимости функционирования нейрона от его метаболизма. Это послужило основанием для поисков долгосрочного памятного следа в стойких изменениях химизма нейроплазмы.
Так как основным субстратом функциональных структур нейрона являются белковые соединения, а их различные модификации задаются матрицами РНК, на которых они синтезируются, то была выдвинута гипотеза о том, что информация, входящая в долгосрочную память, «записывается» структурой полинуклеотидной цепи молекулы (X. Хиден, 1959). По этой гипотезе разные условные потоки импульсов создают различное распределение ионных концентраций вдоль молекулы РНК и вызывают специфические для каждого сигнала перемещения нуклеотидов в их цепи. В результате каждый
В качестве экспериментального подтверждения гипотезы о переопределении нуклеотидов в молекуле РНК как механизме формирования долгосрочной памяти могут служить результаты опытов с крысами, которые обучались добираться к пище, балансируя на проволочном мостике (табл. 4).
Таблица 4. Процентное содержание нуклеотидов в РНК ядра Дейтерса при выработке двигательного навыка у крыс (по X. Хидену, Е. Эльхази)
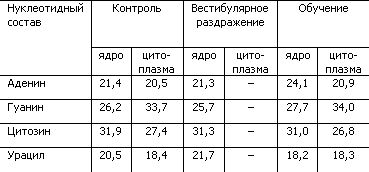 |
Как показывает табл. 4, у крыс, обучившихся сложным движениям, в нервных клетках ядра Дейтерса, ответственного за выполнение нового двигательного навыка, произошло изменение нуклеотидного состава РНК клеточного ядра. Увеличилась доля аденина, уменьшилась доля урацила. Простое вестибулярное раздражение не изменяло нуклеотидный состав. Исследование глиальных клеток ядра Дейтерса выявило сходные изменения РНК, что позволило сделать предположение об участии глии в кодировании поступающих сигналов путем изменения структуры нуклеотидных цепей РНК.
Однако гипотеза о нуклеотидном коде памятного следа встретилась со многими затруднениями, из которых главное —
В связи с этим внимание исследователей обратилось к более долговечным химическим компонентам структурно-функциональной организации нервной клетки. Памятный след стали искать в
Анализ свойств ДНК и влияния ингибиторов ее синтеза на память привел к предположению, что формирование памятного следа связано с активацией определенных генов при деспирализации соответствующих участков цепи ДНК. Это происходит под влиянием ионных сдвигов, отражающих возбуждение нервной клетки при поступлении в нее сигналов.
На основании того, что снятие гистоновой блокады дерепрессирует гены, обусловливающие модификацию свойств клонов клеток была предложена
Электрофизиологические исследования механизмов привыкания нервных клеток к действию раздражителей привели к представлению, что памятный след отражает не столько временные, сколько пространственные характеристики сигналов. Он представляет собой не «паттерн импульсов», а «паттерн синапсов», кодирующих своим расположением и состоянием значение сигнала (Е.Н. Соколов, 1969). При этом медиаторы через рецепторы синаптических мембран вызывают активацию одних и инактивацию других оперонов молекулы ДНК. В результате формируется система обеспечения синтеза специфических белков, реализующих функцию «обученной нервной клетки (рис. 25).
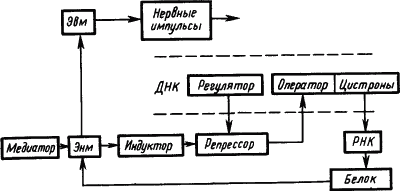 |
Рис. 25. Участие функциональных структур генетического аппарата молекулы ДНК в долговременной памяти (по Е.Н. Соколову):
пунктиром обозначен участок молекулы ДНК; Эвм — электровозбудимая мембрана, Энм — электроневозбудимая мембрана
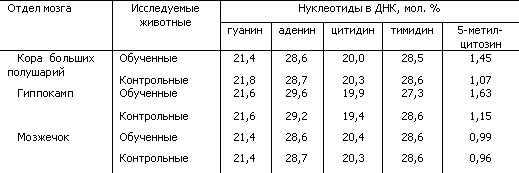
За последнее время получены новые доказательства участия ДНК в механизмах долговременной памяти. Показано, что при выработке условных рефлексов происходят своеобразные изменения структуры молекулы ДНК в нервных клетках мозга крыс. Эти изменения состояли в увеличении степени метилирования ДНК, повышающего активность определенных генов. Они были четко выражены в коре больших полушарий и гиппокампе и оказались незначительными в мозжечке. При такой «мягкой» обратимой модификации ДНК ее нуклеотидный состав оставался неизменным (табл. 5).
Таблица 5. Содержание нуклеотидов и метиловых компонентов в ДНК из разных отделов мозга у обученных и контрольных крыс (по Н.А. Тушмаловой)
 |
Как уже было упомянуто, основные структуры нервной клетки, реализующие ее функции, состоят главным образом из белковых соединений. Поэтому многие исследователи ведут изучение
Исследование белков мозга голубей выявило, что при их обучении происходит возрастание фракции мембранных гликопротеидов, обозначенных 10
Значение белков в механизмах формирования долгосрочной памяти продемонстрировано в опытах, где угнетение синтеза белка нарушало переход краткосрочной памяти в долгосрочную. Так, введение ингибитора белкового синтеза пуромицина золотым рыбкам сразу после обучения стирало его следы, но введение пуромицина через 1 ч не вызывало такого действия. Эти опыты были повторены и на других животных, показывая, что синтез белка необходим для консолидации памяти, а когда памятный след закреплен, белок уже не столь нужен.
Какие же именно белки разрушаются под действием ингибитора, препятствующего формированию долговременной памяти? Для ответа на этот вопрос исследовали разные фракции мембранных белков синаптического аппарата после нарушения долговременной памяти пуромицином. Показано, что этот ингибитор наиболее резко тормозит синтез кислых белков, к которым относится специфический для тканей мозга белок S-100. Оказалось также, что белки наиболее чувствительной к ингибитору фракции по своему аминокислотному составу принадлежат к группе внутренних белков мембраны, которые связаны с ее функциями как холинорецепторы. Это соответствует мнению, что носителями долгосрочной памяти являются холинэргические синапсы. Однако имеются указания на участие в механизмах долгосрочной памяти и адренэргической системы. Отмечено, что положительное подкрепление условных рефлексов, например пищевых, связано с функционированием преимущественно серотонинэргических, а отрицательное при оборонительных рефлексах — норадренэргических структур мозга. О возможных нейрохимических компонентах подкрепления существуют разные мнения: сюда относят и норадренэргическую систему, и дофаминэргическую. За последнее время накоплено большое количество фактов, показывающих, как участвуют нейрохимические механизмы в процессах обучения и памяти.
Другой подход к выяснению роли белков в долговременной памяти состоял в прослеживании включения в них меченых аминокислот и изменений этих белков при обучении. Так, внедрение меченного тритием лейцина в белки мозга крыс после обучения возрастало и сопровождалось увеличением числа рибосом, связанных с мембранами. При переучивании крыс способу добывания пищи на стадии бурной двигательной активности происходило даже снижение включения меченого лейцина в белки мозга. По мере усвоения нового навыка включение меченого лейцина в белки, особенно специфические, возрастает. После выработки нового навыка пlt;...пропуск части слова...gt;ния у золотых рыбок отмечен синтез трех новых фракций белков, обнаруженных с помощью меченого валина. У крыс, обученных дифференцированию зрительных сигналов для решения лабиринтной задачи, наблюдали ускоренное включение меченого тирозина в мембранные белки. Выработка условных рефлексов у голубей сопровождалась усилением синтеза гликопротеинов по показателям включения меченого валина.
В последние годы все более широко применяют методы иммунохимической идентификации специфичных для мозга белков, в частности S-100, 14-3-2; гликопротеинов для определения их роли в фазировании свойств возбудимых мембран и синаптических процессов. Таким образом, накапливаются веские доказательства непосредственного участия белков, особенно кислых мембранных и гликопротеинов в переходе краткосрочной памяти в долгосрочную.
Сравнительно недавно было выяснено, что к возможным носителям долгосрочной памяти относится ряд пептидов — веществ, как и белки, состоящих из аминокислот, но более простых по своему строению.
К этим пептидам относятся главным образом такие гормоны гипофиза, как адренокортикотропин, меланоцитостимулирующий гормон, вазопрессин, окситоцин и некоторые рилизинг-гормоны; а также пептиды, специфически связанные с обучением какому-либо одному определенному навыку. Связь пептидов с памятью, по-видимому, определяется их участием в синаптической передаче. Предполагают даже существование особых «пептидэргических синусов», которые могут изменять свои свойства проведения.
Кроме того, считают, что факторы пептидной природы могут участвовать в образовании временных связей как химический механизм воздействия подкрепляющих систем на генетический аппарат нервных клеток.
Одно из замечательных свойств памяти состоит в специфической избирательности и локальности образуемых временных связей. В хорошо выработанном условном рефлексе каждый сигнал вызывает только свою строго определенную реакцию. В этом, а также в длительности хранения, свойствах угасания и восстановления, в чувствительности к ядам, тормозящим синтез белков и РНК, «память» нервной системы очень напоминает «память» системы иммунитета. Предположение об участии иммунных реакций в формировании долгосрочной памяти получило новые подтверждения и привело к разработке
Согласно этой гипотезе в результате метаболизма синаптических мембран на их поверхности образуются вещества, которые, накапливаясь в избытке, при реверберации импульсов в стадии краткосрочной памяти играют роль антигена для антител, вырабатываемых в глиальных клетках. Связывание антитела с антигеном происходит при участии активаторов образования медиаторов или ингибитора ферментов, расщепляющих эти активаторы (рис. 26). В пользу этой гипотезы свидетельствуют эксперименты, показавшие, что антитела к синаптосомам повышали чувствительность последних к медиаторам и что стимуляторы иммуногенеза улучшали долгосрочную память.
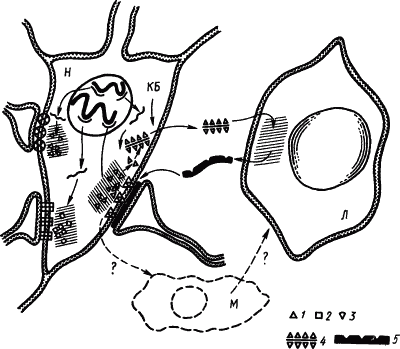 |
Рис. 26. Иммунологические механизмы, повышающие проводимость определенных синапсов (по И.П. Ашмарину):
КБ — катионные белки, открывающие путь комплексам фрагментов антигена синапса с РНК через оболочку клетки; Л — клетки глии, функционально подобные лимфоцитам, М — клетки глии, функционально подобные макрофагам;
Участие в иммунохимических механизмах памяти специфичного для мозга белка S-100 показано в опытах эпидурального введения крысам сыворотки с антителами к этому белку, в результате чего затруднялась выработка навыка поведения в лабиринте. Чтобы выделить в «чистом виде» эффект действия антител на белки, специфические для процессов образования временных связей, изучали навык поведения крыс. При этом вводили антитела, которые вырабатывались на гомогенаты мозга необученных и обученных крыс, а также крыс, получавших препараты, облегчающие или затрудняющие обучение.
Эффект действия антител резко проявлялся уже в 1-й день обучения в виде угнетения условных реакций животных, получавших антитела к веществам-участникам формирования долговременной памяти под влиянием препаратов, облегчающих обучение. Видимо, такой эффект объясняется связыванием и дезактивацией этих веществ.
Больше половины мозга занимают глиальные клетки, которых примерно в 10 раз больше, чем нервных; каждый нейрон со всех сторон окружен глией. Естественным было предположение, что глия участвует в осуществлении функций мозга, в том числе высшей нервной деятельности. Замедленность реакций клеток глии и сосредоточение в них специфичных для мозга белков S-100, некоторых гликопротеинов и биологически активных соединений создают условия для реализации постепенно стабилизирующихся процессов, к каким относится память. Поэтому появились основания предполагать, что
Для подтверждения этого предположения проводили эксперименты, показавшие, что инъекции экстрактов глии крысам ускоряли выработку условных рефлексов. В пользу такого предположения свидетельствует и прогрессивное увеличение глиальной ткани относительно нервной в ряду от низших животных к высшим. Медленная электрическая активность глии может быть источником колебаний постоянного потенциала мозга и электротонических потенциалов, создающих очаги повышенной возбудимости, которые считают основой образования временных связей.
О возможном механизме участия глии в фиксации памятного следа существуют различные мнения. Так, предполагали, что выходящие при возбуждении нейрона ионы K захватываются клетками глии и передаются следующему нейрону, вызывая стойкое изменение уровня его поляризации, а следовательно, облегчение его возбуждения. Другое объяснение состоит в том, что выработка условного рефлекса вызывает у прилегающих к нейрону глиальных клеток усиленное образование миелина, который окутывает его тончайшие терминальные волокна и уменьшает фактор риска проведения по ним импульсов, в результате чего «потенциальные» синапсы становятся действующими (А.И. Ройтбак, 1968). Схема происходящих при этом процессов представлена на рис. 27. Эта гипотеза получила экспериментальное подтверждение в стимуляции миелинообразования глиальными клетками с помощью ионов K, которые выходят из возбужденного нейрона в межклеточную среду.
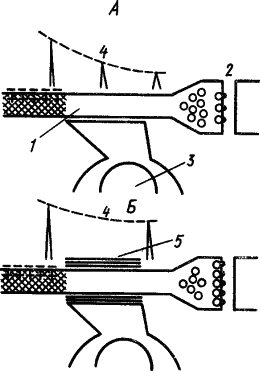 |
Рис. 27. Гипотеза о роли глии в образовании временной связи условного рефлекса.
Для того чтобы определить, какова в долговременной памяти роль глии и нейронов, испытывали действие на условные рефлексы крыс антител, полученных при иммунизации гомогенатом или глии, или нейронов. Оказалось, что антинейрональная сыворотка ухудшала главным образом краткосрочную память, а антиглиальная — долгосрочную. Эти результаты рассматривают как указание на то, что краткосрочная память обусловлена реверберацией импульсов по цепям нейронов, а долгосрочная — реализуется пластическими процессами с участием глии.
Уже давно высказали мнение, что
Качественное своеобразие системной деятельности нейронов обосновывается морфологическими особенностями их связей (Я. Сентаготаи, 1968), эффектами объединения; существует представление о том, что при обучении временные связи формируются путем объединения нейронов различных структур мозга в комплексы, называемые
Существует концепция, по которой каждый акт высшей нервной деятельности связан с активацией определенной совокупности нейронов, образующей сложную мозаику. Такой подход к проблеме памяти лишен затруднений, возникающих перед гипотезами ее «молекулярного кодирования», и подкрепляет концепцию «распределенной памяти», согласно которой
Ha основании большого экспериментального материала о взаимосвязи ритмических свойств нейронов, образующих цепочки возбуждения разной лабильности и определяющих функциональное состояние центральных нервных структур, было высказано мнение, что новая временная связь запечатлевается в нервной системе в виде специфического пространственно-временного распределения активности нейронов, перестроивших свою лабильность (М.Н. Ливанов, 1975). В пользу этого предположения свидетельствуют результаты исследования конфигурации мозаики нейронной активности, показавшие, что условный рефлекс проявляется лишь тогда, когда рисунок мозаики, вызванной сигналом, соответствует таковому памятных следов и подходит к нему, как «ключ к замку». Возникновение разных рисунков нейронной активности с участием одних и тех же нейронов означает перераспределение свойств проводимости их синапсов, которое может осуществляться рассмотренными выше механизмами.
Такое представление близко к взглядам на нейронную структуру условного рефлекса, как на массовое объединение нейронов, в котором воспроизводится вся ситуация в целом (А. Аттли, Т. Блумфилд, Е. Джон, 1973). Обобщение большого материала многолетних нейрохимических исследований памяти также привело к заключению о ее распределенном характере в ансамблях нейронов. При этом роль биохимических процессов, создающих сложные «рисунки» распределения химизма нервных клеток, заключается в организации «узнавания» клетками друг друга и «сборки» их в функциональные ансамбли. Существует предположение о химической природе сигнала, управляющего проходимостью синапсов нейронов. Это выражается перестройкой классов доминирующих межимпульсных интервалов, в активности нейронов.
Получены также факты, указывающие на то, что механизм образования условного рефлекса не ограничивается перестройками центральных процессов, а включает в себя и
Особое место в исследованиях занимают явления, получившие название
Результаты экспериментов по изучению условных реакций планарий, получавших экстракт РНК от «обученных» и «необученных» особей, дали основание предполагать, что «переносчиком» памяти может служить именно РНК.
Опыты проводились и на позвоночных животных, которым вводили экстракт мозга «обученных» животных. Так, экстракт мозга рыбок, у которых был выработан условный рефлекс избегания ударов тока по сигналам синего или зеленого цвета, при введении его необученным рыбкам вызывал у них отрицательную реакцию на соответствующий цвет. Многочисленные опыты с «переносом» памяти были поставлены на крысах, причем были получены как положительные, так и отрицательные результаты (например, исследование влияния такого экстракта мозга на выработку целевых условных рефлексов).
Поиски активного начала мозговых экстрактов привели к выделению из них пептидов, обнаруживших высокую избирательность своего действия (Г. Унгар, 1972). Так, из экстракта мозга крыс, обученных избегать темноту (его введение вызывало у необученных также избегание темноты), был выделен пептид, состоящий из 15 аминокислот. Он получил название
Исходя из предположения о том, что нейроны «узнают» друг друга по химическим меткам (в частности, пептидной природы), была высказана гипотеза о формировании на этой основе памятных следов (рис. 28). На рис. 28 показано, как пептид
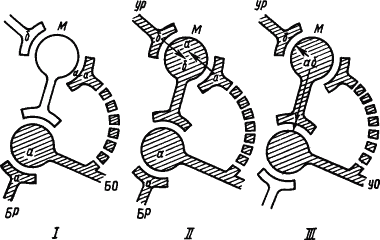 |
Рис. 28. Пептидный механизм образования новых путей проведения импульсов.
БР — раздражитель безусловного рефлекса, БО — безусловный ответ, УР — условный раздражитель, УО — условный ответ, М — «клетка памяти»,
Описано возникновение реакции вкусового отвращения к сахарину у крыс, получавших гомогенизированный экстракт мозга от крыс, у которых эта реакция вырабатывалась сочетанием питья раствора сахарина с действием ионизирующей, радиации. Аналогичные результаты получены при другом подкреплении: если у собак с односторонними слюнными условными рефлексами сделать частичный обмен цереброспинальной жидкостью, то наступают изменения рефлексов, появляющиеся на «донорской» стороне. Отмечено, что выработка условного рефлекса отказа от пищи у виноградной улитки происходит быстрее, если ей перелить гемолимфу из цефалопедального синуса ранее обученной улитки.
Однако концепция переноса памяти не является общепризнанной. Сказываются сомнения как по поводу применяемых методик и достоверности результатов, так и в отношении их трактовки. Так, например, остается неясным, как может «переноситься» программа сложного акта поведения и каков должен быть механизм ее реализации. Вопрос о влиянии экстрактов мозга обученных животных на поведение получивших его необученных, несомненно, имеет большое значение для познания механизмов памяти, но требует дальнейших исследований.
| © 2025 Библиотека RealLib.org (support [a t] reallib.org) |