"Основы физиологии высшей нервной деятельности" - читать интересную книгу автора (Коган Александр Борисович)
Глава 4 ТОРМОЖЕНИЕ УСЛОВНЫХ РЕФЛЕКСОВ
До сих пор был рассмотрен лишь один основной физиологический процесс высшей нервной деятельности: процесс
Торможение — главнейшее средство упорядочения и совершенствования условных рефлексов. Благодаря торможению достигается сосредоточенность на наиболее важной в данный момент для организма деятельности и задерживается все второстепенное
Взаимодействие всех этих видов торможения обеспечивает замечательную точность и гибкость высшей нервной деятельности.
В процессе жизнедеятельности организм постоянно подвергается тому или иному раздражению извне или изнутри. Каждое этих раздражений способно вызвать соответствующий рефлекс. Если бы все эти рефлексы могли реализоваться, то деятельность организма была бы хаотической. Однако этого не происходит. Наоборот, для рефлекторной деятельности характерны согласованность и упорядоченность: с помощью безусловного торможения наиболее важный в данный момент для организма рефлекс на время своего осуществления задерживает все остальные, второстепенные рефлексы.
Безусловное торможение свойственно всем элементам нервной системы. Его не нужно вырабатывать, оно сразу появляется с началом рефлекса, вызывающего угнетение остальных рефлексов, и исчезает после его прекращения. Безусловное торможение называют также
Постоянный тормоз. У собаки с хорошо выработанным условным слюнным рефлексом каждое применение сигнала вызывает обильное выделение слюны. Однако если включить сигнал и одновременно причинить собаке боль, то из ее слюнного протока не выделится ни капли слюны: условный рефлекс исчез. Угнетение условного слюноотделительного рефлекса произошло вследствие безусловного торможения, связанного с возникновением жизненно важного для организма оборонительного рефлекса. Вредящее раздражение — настолько значительное событие, что оно требует сосредоточения всех усилий организма на рефлексе защиты от раздражающего воздействия. Безусловное торможение и обеспечивает такое сосредоточение, задерживая все другие рефлексы.
Можно повторно много раз наносить собаке болевое раздражение, и каждый раз она будет на него реагировать. В соответствии с постоянным характером безусловных рефлексов и связанное с ними безусловное торможение проявляется постоянно, не ослабевая при повторениях.
Источником постоянного безусловного торможения могут быть разнообразные безусловные рефлексы, часто это бывают рефлексы внутренних органов. Например, во время опыта у собаки без всякой видимой причины исчезают все условные рефлексы. Но стоит вывести ее на улицу и освободить мочевой пузырь, как все рефлексы немедленно восстанавливаются: они были заторможены импульсами от интерорецепторов растянутого мочевого пузыря.
Жизнь животных в природе дает множество примеров безусловного постоянного торможения условных рефлексов. Пожирая добычу, хищник становится менее осторожным, так как безусловный пищевой рефлекс тормозит условные оборонительные. О силе торможения, вызванного болевым рефлексом на все остальные рефлексы, можно судить по тому, что раненый зверь «забывает» воспитанный всей жизнью страх перед человеком и может броситься на охотника.
Торможение высшей нервной деятельности человека безусловными рефлексами является повседневным фактом. Тормозящее действие пищеварительных рефлексов, в частности, от интерорецепторов желудка известно каждому. При сильной боли человек теряет способность спокойно рассуждать и «забывает все на свете». Говорят, что у него «все заслонила боль». Так сильно проявляется безусловное торможение, связанное с защитным рефлексом.
Источником безусловного торможения условного рефлекса может также стать другой условный рефлекс. Торможение одних условных рефлексов другими придает особую гибкость поведению животных. Привлекаемая видом зерна, рассыпанного в ловушке, птица подходит все ближе к силкам. Но лишь треснет сухая ветка под ногой неосторожного ловца, как птица взвивается в воздух. Реакция бегства как условный оборонительный рефлекс мгновенно затормозила условный пищевой рефлекс. Условный половой рефлекс у лошади может полностью затормозить условные рефлексы, необходимые при езде.
Человек, обладающий разнообразнейшими условными связями, имеет очень много поводов для торможения одних видов своей деятельности другими. Каждому известно, как одно впечатление может подавлять другие. Например, если мы хотим вслушаться в тихую речь, то закрываем глаза, чтобы прекратить действие зрительных раздражений. Но наиболее важное место занимают в мышлении и поведении человека те случаи, которые психология обозначает как «волевую деятельность». Так, условный оборонительный рефлекс исключительной биологической силы заставляет солдата, залегшего под обстрелом, прижиматься к земле (речь идет о жизни или смерти). Казалось бы, он весь во власти защитной реакции. Но раздается команда: «За Родину, вперед!», рядом поднимаются товарищи, и только что непреодолимый условный оборонительный рефлекс затормаживается высшими условными связями сознания. Человек переживает этот процесс в его субъективном выражении, как подавление животного страха сознательным чувством долга.
Гаснущий тормоз. Безусловное торможение условных рефлексов вызывается также часто возникающим ориентировочным рефлексом. Например, если во время опыта с условным слюноотделением слегка постучать по стене, то собака немедленно насторожится, а условное слюноотделение прекратится. Однако если такое постукивание повторяют многократно, то реакция на постукивание постепенно ослабевает, слюноотделение тормозится все меньше и, наконец, совсем растормаживается. Часто повторяемый ориентировочный рефлекс становится привычным и теряет свое тормозящее действие.
В соответствии с вре́менным характером ориентировочного рефлекса связанное с ним безусловное торможение постепенно ослабевает при повторении, поэтому оно и называется
Тот факт, что ориентировочный рефлекс в однообразной, не меняющейся обстановке постепенно исчезает, имеет большой биологический смысл.
Например, поведение котенка, впервые попавшего во двор: он замирает перед хлопающим крыльями петухом, бросается в сторону от мычащей коровы, застывает, принюхиваясь к запаху из конюшни. Все для него ново, за каждым незнакомым предметом, звуком, запахом может скрываться неведомая опасность. Однако если бы эти ориентировочные реакции оставались на всю жизнь, они затормозили бы всю высшую нервную деятельность животного. Постоянно настороженное животное не могло бы образовывать новые условные рефлексы, не могло бы ничему научиться.
Так не бывает, не бывает именно потому, что ориентировочные рефлексы и связанное с ними торможение условных рефлексов при повторениях постепенно ослабевают и, наконец, полностью исчезают. Поэтому вскоре обстановка двора перестает вызывать на себя ориентировочные реакции и не мешает котенку приобретать необходимый жизненный опыт.
Человек постоянно испытывает действие гаснущего тормоза. Школьникам, особенно малышам, всегда нужно дать «оглядеться» в классе, чтобы ориентировочные рефлексы угасли и не мешали усвоению урока. В незнакомой аудитории лектор иногда теряет lt;...слово не видно в скане...gt; мысли и может вдруг забыть хорошо известные ему понятия, однако, немного привыкнув к новым слушателям, он легко вспоминает даже мельчайшие подробности.
Высшая нервная деятельность со временем уточняется — поведение избавляется от всего ненужного, устаревшего, всесторонне совершенствуется. Все это связано с тормозными процессами, развиваемыми каждым условным рефлексом в отношении самого себя. Благодаря этому рефлексы отменяются, если сигналы потеряли сигнальное значение
Для всех этих случаев торможения общим является то, что оно вырабатывается, т.е. является
Угасательное торможение. Если собаке с хорошо выработанным условным пищевым слюноотделительным рефлексом подавать условный раздражитель, не сопровождая его подкармливанием, то слюны будет выделяться все меньше, а само выделение будет наступать со все большим латентным периодом (табл. 6).
Таблица 6. Уменьшение условного слюноотделения на метроном при его применении без пищевых подкреплений (опыт на лекции И.П. Павлова, 1927)
Порядковый номер применения сигнала без подкрепления // люноотделение за 30 с действия сигнала, капли // Время от начала слюноотделения (латентный период), с
1 // 10 // 3
2 // 7 // 7
3 // 8 // 5
4 // 5 // 4
5 // 7 // 5
6 // 4 // 9
7 // 3 // 13
Отсутствие подкрепления рано или поздно приводит к тому, что сигнал перестает совсем оказывать влияние на слюнную железу: условный рефлекс исчезает.
Таким образом,
Угасательное торможение представляет собой очень распространенное явление и имеет большое биологическое значение. Нетрудно представить себе, что было бы, если бы все образовавшиеся в разное время условные рефлексы, переставшие соответствовать новым условиям, оставались на всю жизнь. Тогда проголодавшаяся собака долго бесполезно бегала бы «по старой памяти», например, к пустому дому, жильцы которого когда-то ее покормили. Или вышедшая на охоту лисица бесконечно сидела бы у какой-нибудь пустой сусличьей норы. Угашение условного рефлекса происходит тем быстрее, чем чаще предъявляется условный раздражитель без подкрепления (рис. 29).
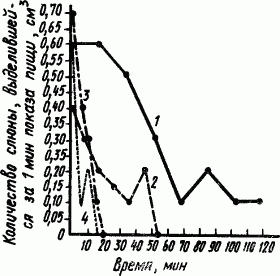 |
Рис. 29. Угашение натурального пищевого слюноотделительного рефлекса при разной частоте показа мясного порошка (без скармливания) (по Б.П. Бабкину):
От устаревших и бесполезных условных связей высшая нервная деятельность освобождается путем их угашения. Люди расстаются с устаревшими взглядами, когда оказывается, что идущая вперед жизнь не подкрепляет ранее сложившихся у них представлений.
Дифференцировочное торможение. У собаки выработали условный рефлекс пищевого слюноотделения на стук метронома 60 ударов/мин. Каждое включение метронома с такой частотой вызывает условное слюноотделение. Но если применить другой метроном с частотой 120 ударов/мин, то и он тоже вызовет отделение слюны, хотя сам никогда не сочетался с едой.
Однако достаточно несколько раз не подкрепить действие метронома с частотой 120 ударов/мин, как он перестает быть сигналом еды и наступает дифференцирование этого раздражителя от метронома с частотой 60 ударов/мин, сопровождающегося едой. Теперь метроном с частотой 120 ударов/мин никакого слюноотделения уже не вызывает, что является результатом дифференцировочного торможения.
Насколько важное место в жизни занимает дифференцировочное торможение, видно, если, например, сравнить поведение неопытного щенка с поведением взрослой собаки, выработавшей дифференцировки.
Хотя щенка кормит хозяин, он подбегает и к чужим людям. Обратив в бегство воробьев, он начинает гонять по двору кур. Каким контрастом с постоянными ошибками и промахами неопытного щенка выглядит поведение взрослой собаки, умеющей дифференцировать даже разные интонации голоса своего хозяина. Услышав ласковые нотки, она подбегает к нему, а когда в голосе хозяина звучит раздражение, уходит подальше.
Непрерывное, все более тонкое различение, дифференцирование явлений окружающего мира составляет важную часть мышления человека. Человеческий ум переносит центр тяжести такого различения в область понятий, выраженных речью. Путем дифференцирования словесных раздражителей выявляются их частные особенности, необходимые для образования новых понятий.
Условный тормоз. Если при хорошо укрепленном условном слюнном рефлексе, выработанном на стук метронома, время от времени применять метроном вместе с дополнительным раздражителем, например бульканьем, не сопровождая эту комбинацию пищевым подкреплением, то вскоре сам метроном по-прежнему будет вызывать отделение слюны, а в сочетании с бульканьем потеряет свое положительное действие. Прибавочный агент стал условным тормозом.
Образуемые жизнью условные тормозы уточняют условные рефлексы и имеют большое приспособительное значение в поведении животных.
Так, издалека заметив ползущую гусеницу, цыпленок бросается к ней, но, подбежав, видит длинные волоски — примету несъедобности — и отходит в сторону. Вид волосков — это прибавочный агент, отменяющий пищевое значение условного раздражителя. Когда в лесу слышится шуршание сухих листьев, то притаившийся под кустом заяц настораживается, готовясь к бегству. Но порыв ветра доносит успокаивающий запах безобидного ежа, и заяц снова принимается за обгладывание веток. Жизненный опыт сделал этот запах условным тормозом оборонительного рефлекса бегства от звуков, обычно сигнализирующих приближение опасности.
Торможение запаздывания. Если у собаки с прочно выработанным совпадающим слюнным условным рефлексом увеличивать изолированное действие условного раздражителя до 1, 2 и даже 3 мин, считая от момента включения сигнала до момента подкрепления, то слюноотделение начнет все более «запаздывать». Возникновение условного слюноотделения будет постепенно приближаться к моменту подкармливания. Большая же часть времени действия раздражителя с момента его включения является недеятельной фазой условного рефлекса, и она связана с развитием торможения запаздывания.
Таким образом, один и тот же условный раздражитель сначала действует тормозящим образом, а потом возбуждающим. Этот парадокс объясняется тем, что появляется другой раздражитель —
Хорошим примером приспособительного значения торможения запаздывания может служить условное выделение желудочного сока. Благодаря торможению запаздывания, избегается бесполезное, а возможно, и вредное наполнение пустого желудка кислым желудочным соком, «запальный» сок вовремя встречает поступающую в желудок пищу, обеспечивая полноценное ее переваривание.
Торможение возникает в каждое мгновение нервной деятельности. Выше были рассмотрены основные его виды, каждый в отдельности, в связи с причинами их возникновения. Однако в жизни поводы для возникновения торможения так переплетаются, что развитие торможения всегда происходит в условиях сложного взаимодействия разных его видов. Можно наметить два главных типа такого взаимодействия: взаимодействие раздражителей, когда один тормозной процесс ослабляет или даже уничтожает другой —
Растормаживание угашенного условного рефлекса. Показывая студентам опыт с угасанием условного слюноотделительного рефлекса, И.П. Павлов обратил внимание слушателей на следующую подробность. Как правило, с каждым применением сигнала без подкрепления величина рефлекса становилась все меньше. Но неожиданно при очередном неподкреплении слюна вдруг пошла быстрее. Это совпало с небольшим шумом в аудитории, т.е. внешним торможением. Потом опять наступила тишина и угасание рефлекса продолжалось вновь. Следовательно, внешнее торможение, натолкнувшись на внутреннее, может его ослабить. Однако это ослабление прекращается, как только прекратится действие внешней причины. Действительно, уже следующее применение неподкрепленного сигнала при установившейся в аудитории тишине дало вновь резкое падение величины угашаемого рефлекса.
Взаимодействие внешнего и внутреннего торможения в этих случаях выступает в наиболее простой форме. Внешнее торможение, встретившись с внутренним, уничтожает его. Одно торможение тормозит другое, и, образно говоря, два отрицательных явления приводят к положительному результату. В этом и состоит внешнее проявление растормаживания. Растормаживание угашенного рефлекса заключается во временном устранении угасательного торможения при помощи внешнего. Оно вызывается разнообразными видами внешнего тормоза (безусловными и условными рефлексами) умеренной силы и проявляется в полном или частичном восстановлении рефлекса во время их действия.
Растормаживание дифференцировок. Растормаживание дифференцировок наглядно демонстрирует следующий опыт. У собаки с условным пищевым рефлексом слюноотделения на звук тона было выработано дифференцирование неподкрепляемого пищей звука 1/8 тона. В многочисленных пробах тон положительный сигнальный постоянно вызывал отделение слюны, а применение 1/8 тона такого действия не имело. Затем были взяты посторонние раздражители — звук бульканья и запах амилового эфира. Испытанные каждый по отдельности, они не только не вызывали отделения слюны, но в качестве гаснущего тормоза даже временно задерживали положительные слюноотделительные рефлексы. Однако если присоединить один из этих видов тормоза к тоже тормозному дифференцировочному раздражителю 1/8 тона, то последний растормаживается. Он превращается в положительный раздражитель и вызывает отделение слюны, пока действует внешний тормоз.
Растормаживание дифференцировок заключается в срочном устранении дифференцировочного торможения внешним. Оно происходит под влиянием внешнего тормоза умеренной силы и проявляется во временном возвращении дифференцировочному раздражителю его положительного условного действия. В результате теряется выработанное различение подкрепляемых и неподкрепляемых разновидностей сигнала.
Растормаживание условного тормоза. Условный тормоз, как и другие виды внутреннего торможения, при встрече с внешним торможением разрушается. Это показывает, например, опыт с действием таких посторонних агентов, как метроном или тепловое раздражение кожи, на условно-тормозную комбинацию из вертящегося предмета (положительный сигнал) и тона (прибавочный отрицательный агент).
Прочно выработанная условно-тормозная комбинация вида вертящегося предмета со звуком тона при добавлении постороннего раздражителя (метронома) сразу потеряла свое тормозное значение и вызвала отделение слюны. Но повторное испытание тормозной комбинации без присоединения метронома показало, что уже через несколько минут на нее опять не выделяется слюна. Растормаживание бесследно исчезло. Следовательно,
Так же, как и в других случаях растормаживания, гаснущие тормозы при повторных применениях действуют все слабее. С каждым разом уменьшается вызываемая ими ориентировочная реакция и все меньший ущерб наносится внутреннему торможению. В то же время безусловные и условные постоянные тормозы могут стойко препятствовать отрицательному действию условно-тормозной комбинации.
Растормаживание запаздывающего условного рефлекса. Когда у собаки хорошо выработано запаздывание условного слюнного рефлекса, то при включении условного раздражителя собака не будет на него реагировать, пока не приблизится срок подкрепления. В течение одной, двух и даже трех минут непрерывного действия сигнала слюна не выделяется. Однако если в это время к бездействующему сигналу присоединить постороннее раздражение, то слюна потечет немедленно. Прорвется сдерживаемый торможением запаздывания условный рефлекс, но к моменту подкрепления он окажется подавленным. В этом факте открывается двоякое действие постороннего агента: растормаживание недеятельной части сигнала и затормаживание деятельной. Внешнее торможение в данных условиях опыта освобождает задержанный рефлекс от условного торможения, но, заставая рефлекс готовым к действию, само его задерживает. В других условиях опыта, при иных временных и силовых отношениях постороннего агента и сигнала, результаты будут другими.
Таким образом,
Растормаживание изменяет приспособительное значение условных рефлексов, аннулируя уточнение условного рефлекса, определенно снижает его приспособительную ценность. Но вместе с тем в чрезвычайных обстоятельствах (а это и есть обстановка действия внешнего тормоза) возврат к грубым, неуточненным, зато более универсальным формам рефлексов может иметь положительное значение.
Например, у лисицы, которая повадилась таскать кур с птицефермы, выработалось дифференцирование: она скрывалась при виде собаки, а при виде сторожа, который не мог ее догнать, только отбегала подальше. Надеясь подстрелить похитительницу, сторож обзаводится ружьем, но теперь лисица скрывается и от него: вид необычного предмета — ружья — подействовал как внешний тормоз и вернул лисице ее былую осторожность, растормозив оборонительный рефлекс бегства.
Каждый знает, какое воздействие может оказать такой внешний тормоз, как неожиданное известие или необычайное событие. При этом человек начинает, путать то, что раньше различал (растормаживание дифференцировок). Вместе с тем в этой ситуации он может вспомнить давно забытое (растормаживание угашенных связей).
Суммация торможения. Встреча тормозных процессов далеко не всегда приводит к их ущербу, как это было в случае растормаживания. В результате такой встречи может произойти их сложение и усиление. Наиболее часто такое сложение, или
Можно наглядно показать суммацию двух разновидностей внутреннего торможения, например угасательного и запаздывательного. Для этого у собаки производят угашение ранее выработанных условных рефлексов. Один раз угашают короткоотставленный рефлекс, т.е. практически вырабатывают угасательное торможение. Второй раз, для сравнения, угашают рефлекс после его перевода на длительное отставление, т.е. одновременно вырабатывают угасательное и запаздывательное торможение. Оказалось, что рефлекс угасает быстрее во втором случае (рис. 30). Угасательное торможение с помощью запаздывательного полностью задерживало рефлекс после двух неподкреплений. Без запаздывательного оно подавляло полностью рефлекс только после 16 неподкреплений.
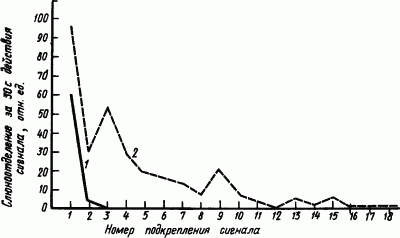 |
Рис. 30. Угашение условного пищевого рефлекса слюноотделения на свет при разном отставлении неподкрепляемого условного раздражения:
При соответствующих условиях может суммироваться даже внешнее торможение с внутренним.
Рассмотрение разновидностей внутреннего торможения: угасательного, дифференцировочного, условного и запаздывательного — показывает, что все они — разные поводы к развитию одного и того же нервного процесса. Отсюда неудивительна их способность к суммации. Факты суммирования внешнего торможения с внутренним, переход одного из них в другое показывают, что даже внешнее и внутреннее — разные виды торможения — имеют одну физиологическую природу.
После того как И.М. Сеченов открыл центральное торможение, было много попыток понять, как может одна нервная клетка привести другую в недеятельное состояние. Однако до сих пор природа торможения остается до конца нерешенным и, как говорил И.П. Павлов, «проклятым» вопросом физиологии. Не разбирая громадный материал часто противоречивых сведений о тормозных процессах в разных отделах нервной системы, необходимо сосредоточить внимание на торможении, свойственном ее высшим отделам, — на
Условное торможение как активный процесс. Активный характер условного внутреннего торможения заключается уже в том, что оно вырабатывается нервными клетками. Внутреннее торможение представляет собой результат деятельности нервных клеток и нередко, как, например, в случаях тонких дифференцировок, очень сложной деятельности. Однако торможение не является утомлением, поскольку в таком случае всякое ослабление нервных клеток, например наркотиками, ускоряло бы и усиливало дифференцирование, запаздывание и другие формы условного торможения. В действительности же происходит наоборот: ослабление корковых клеток устраняет торможение. Очень убедительным доказательством активного характера условного торможения является факт растормаживания отрицательных рефлексов дополнительными раздражениями. Торможение снимается дополнительной работой клетки. Если бы торможение было утомлением, то каждое новое усилие могло его только углублять, но никак не устранять.
Активная природа отдельных разновидностей условного торможения обнаруживается в основных свойствах их проявлений. Например, у собаки были выработаны два условных слюнных рефлекса: на стук метронома и звук звонка. Если угашать рефлекс на стук метронома, а затем испытать рефлекс на звонок, то последний окажется уменьшенным. Угасательное торможение, возникшее lt;...пропущено слово...gt; первом рефлексе, распространилось на второй. Но ведь второй рефлекс «не работал» и в нем не может быть утомления. Следовательно, угасательное торможение не является результатом истощения нейронов.
Также не может быть следствием истощения запаздывательное торможение, так как по мере его выработки величина рефлекса растет, т.е. усиливается развиваемое клеткой условное возбуждение. Несовместимы с утомлением и свойства других разновидностей внутреннего торможения.
Но если внутреннее торможение является активным процессом, тяжелой работой, то как с этим примирить его восстановительное действие (например, в случае сна)? На первый взгляд, здесь явное противоречие. Однако противоречие исчезает, если вспомнить, что условное торможение
Естественно, что отношения работы и отдыха при разных условиях развития торможения могут быть очень различными. Следует также иметь в виду, что при любом виде торможения лишь часть нервных клеток снижает свою импульсную активность, а другая часть ее увеличивает.
Локализация условного торможения. В каких анатомических отделах нервной системы может возникать и развиваться условное торможение? Ясный ответ на этот вопрос дает следующий факт. При повторении индифферентного раздражения у собаки происходит угашение ориентировочного рефлекса на этот раздражитель. Так как это торможение вырабатывается, следовательно, оно является внутренним. Если же проделать такой опыт с собакой, у которой целиком удалены большие полушария мозга, то угашения ориентировочного рефлекса не происходит. Без высших отделов мозга развитие угасательного, как и других форм приобретаемого торможения, становится невозможным. Поэтому безусловные рефлексы и протекают у бесполушарных животных так стереотипно. Осуществляющие их низшие отделы головного и спинного мозга сами не способны к выработке условных форм торможения, хотя и обнаруживают более простые свойства «привыкания». Однако они участвуют в его формировании путем воздействия на функциональное состояние коры главным образом через тормозящую таламокортикальную систему.
Но высшие отделы мозга, в частности кора больших полушарий, имеют сложную структуру. Условный рефлекс образуется благодаря временной связи между анализаторами сигнального раздражения и механизмом подкрепляющей реакции; в каком же звене нейронных сетей, образующих пути осуществления условного рефлекса, вырабатывается условное торможение?
Результаты специальных исследований по изучению локализации условного торможения в корковых структурах мозга приводили ученых к разным взглядам (схематически представлены на рис. 31).
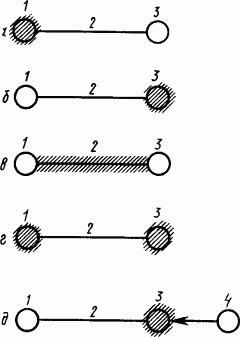 |
Рис. 31. Локализация внутреннего торможения (заштриховано) по представлениям различных авторов (по Л.Г. Воронину):
Первые исследования процесса угашения условного рефлекса, показавшие, что сигнальный стимул при определенных условиях перестает быть действующим, дали основание предполагать, что
Исследование взаимодействия условных слюнных рефлексов при их выработке на базе безусловных, вызываемых раздражением выведенных наружу лоскутков слизистой языка, привело к заключению, что
Систематические исследования функционального состояния всех звеньев структуры условного рефлекса при его угашении привели к заключению, что ни в одном из них не возникал очаг торможения, который мог блокировать условное возбуждение, и оно в зоне проекции сигнального раздражителя становилось источником внутреннего торможения. Этому соответствует тот факт, что нейронные реакции коры мозга кошки при выработке дифференцировочного торможения оказались различными в зонах условного и безусловного раздражителей.
Разногласия по вопросу о том, в каком звене механизма условного рефлекса развивается условное торможение, крайне затрудняют суждение о его нейронной структуре. Однако можно с большой долей вероятности принять, что оно протекает в тех же нервных клетках, которые в свое время участвовали в образовании временной связи. Об этом свидетельствует, к примеру, такой факт, что при угасательном торможении корковый нейрон замедляет свой разряд с тем же латентным периодом, с которым он учащал разряд при положительном рефлексе. В опытах с клеточным аналогом временной связи выработка дифференцировочного торможения изменяла лишь ту часть импульсной реакции нейрона, которая составляла его условный ответ. В процессе развития внутреннего торможения существенную роль играет взаимодействие корковых элементов с подкорковыми, в частности со структурами неспецифической системы таламуса и ретикулярной формации среднего мозга.
Электрофизиологические показатели условного торможения. Когда условный рефлекс угашается, он просто прекращается. В этом является конечный результат деятельности торможения, но не сама деятельность. Некоторые стороны этой деятельности, скрытой от внешнего наблюдения, находят свое отражение в динамике
По мере дальнейшего развития дифференцировочного торможения неподкрепляемый сигнал при каждом своем применении будет усиливать колебания основного ритма и подавлять более быстрые колебания. При этом колебания основного ритма сами сильно замедлятся. Когда дифференцировка будет прочно выработана, отрицательный раздражитель — метроном с частотой 120 ударов/мин — не вызовет никакой двигательной реакции, но каждое его применение станет приводить к изменению электрической активности коры в сторону ее замедления. Часто при этом развивается дремотное состояние и возникают гигантские волны дельта-ритма. Однако медленные волны, наблюдаемые при угашении и выработке дифференцировок, рассматривают как необязательно связанные с развитием сна, а отражающие состояние неподвижности при бодрствовании, обозначая их «ритмами бодрствования — неподвижности». Отмечают, что при глубоком угашении условного рефлекса и выработке прочных дифференцировок медленные волны могут уменьшаться и электрическая активность возвращается к исходному состоянию.
Возможно, переделка ритмических свойств, т.е. подвижности корковых нейронов, составляет наиболее трудную работу торможения. Что касается гигантских размеров медленных волн, то они могут указывать на вовлечение в общий ритм «холостого хода» громадного числа нейронов, освобожденных торможением от разночастотных рабочих возбуждений. Медленные волны возникали при выработке запаздывающего условного рефлекса, при дифференцировании сигналов и во всех других случаях развития внутреннего торможения, что свидетельствует об их общей природе. Однако существуют указания, что в некоторых случаях тормозной раздражитель может не вызвать медленные волны, особенно в начале его действия.
Так как условные связи возникают в результате согласования пространственно-временной организации ритмической активности структур мозга (М.Н. Ливанов, 1970), то при торможении условного рефлекса происходит их рассогласование. Сходство электрических реакций коры на действие положительных и тормозных условных раздражителей привело к заключению, что на уровне коры и тот и другой вызывают процессы возбуждения, а торможение по их командам реализуется на уровне подкорковых исполнительных механизмов.
Таким образом,
Описаны изменения, которые претерпевали при развитии внутреннего торможения
Вместе с тем накапливались факты, свидетельствующие о том, что первичные ответы, отражая общее состояние и уровень возбудимости коры, мало меняются при условно-рефлекторной деятельности. Поэтому особое внимание привлекли поздние компоненты вызванных потенциалов, которые считают наиболее близко отражающими процессы высшей нервной деятельности. Большинство авторов описывают их увеличение.
Из рис. 32 видны наступающее при углублении дифференцировочного торможения возрастание амплитуды первичного ответа и радикальная перестройка поздних компонентов вызванного потенциала. Многие авторы рассматривают усиление поздней поверхностно неlt;...часть слова пропущена...gt;тивной волны как дипольное отражение развития тормозных процессов в глубоких слоях коры. Уменьшение этих волн при подкреплении считают результатом блокирования тормозными клетками импульсации от ретикулярной формации среднего мозга. Однако известно и уменьшение поздних волн вызванных потенциалов. При развитии условного торможения изменяется не только амплитуда, но и форма поздних компонентов вызванных потенциалов, происходит перестройка положительных и отрицательных фаз и появятся новые компоненты.
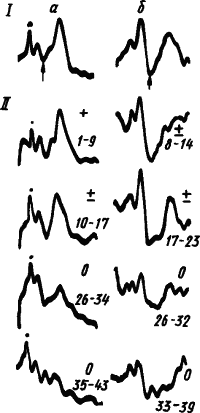 |
Рис. 32. Динамика вызванных потенциалов корковой проекции мышцы смыкания век на щелчки, подкрепляемые обдуванием глаза, при угашении условного рефлекса (по У.Г. Гасанову):
К сожалению, пока не представляется возможным обобщить накопленные весьма разнородные сведения и дать им единое толкование. Видимо, неоднозначность результатов, особенно изменений поздних компонентов вызванных потенциалов, зависит от разных условий эксперимента: влияние посторонних раздражителей, локализации отведения, стадий развития условного торможения, фона электрокортикограммы, двигательной активности животного и многих других условий. В сложной картине динамики поздних негативных и позитивных компонентов вызванных потенциалов отражается развитие активности и смена состояния популяций нейронов разных слоев коры. Не менее сложные отношения складываются при регистрации вызванных потенциалов подкорковых структур. Вместе с тем систематический анализ динамики вызванных потенциалов, особенно его поздних волн, в разных структурах мозга при различных видах внутреннего торможения может быть ценным источником сведений о нейрофизиологическом механизме торможения.
Таким образом,
Так же как образование временной связи условного рефлекса происходило путем совместной деятельности возбуждающихся и тормозящихся нейронов, так и
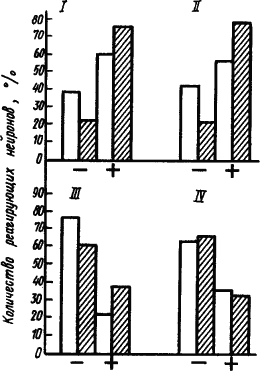 |
Рис. 33. Реакция нейронов коры мозга обезьяны, отвечающих возбуждением (+) или торможением (−) на действие положительного сигнала с электрокожным подкреплением (заштрихованные столбики) и дифференцировочным без подкрепления (белые светлые столбики) (по Г. Джасперу и др.):
Развитие внутреннего торможения, например, при угашении условных рефлексов проявлялось в том, что у нейронов, реагирующих возбуждением, наблюдали снижение частоты условного разряда и даже замедление фоновой импульсации. У нейронов, реагирующих торможением, отмечали повышение частоты разряда на отрицательный сигнал. Наглядная иллюстрация такой динамики нейронных реакций представлена на рис. 34.
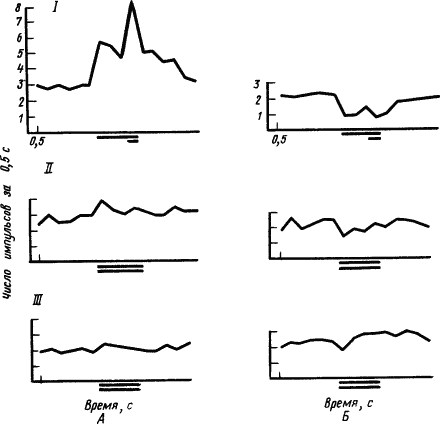 |
Рис. 34. Динамика реакции
длинной линией под абсциссой обозначено время действия условного раздражителя, короткой линией под ней — lt;...gt; безусловного, двумя длинными линиями — действие условного раздражителя без безусловного
Одна из особенностей тормозных реакций нейронов состоит в том, что они угашаются быстрее, чем возбудительные. Так, в опытах с угашением условных рефлексов показано, что в то время как условные реакции возбуждающихся нейронов угашались постепенно и не полностью, реакции тормозящихся нейронов быстро исчезали. При этом наиболее устойчивыми оказывались те фазы импульсного ответа нейрона, которые соответствовали времени бывшего подкрепления. Отмечены существенные различия реакций нейронов разных отделов мозга на развитие внутреннего торможения. Например, нейроны ретикулярной формации среднего мозга крысы при неподкреплении сигнала пищевого рефлекса очень быстро перестают него реагировать, а нейроны гиппокампа еще продолжают отвечать, лишь уменьшив частоту импульсов. Указывают, что импульсные ответы нейронов гиппокампа обладают более высокой точностью дифференцирования по сравнению с другими подкорковыми структурами. Особенности динамики нейронных реакций при выработке условного торможения описаны при исследовании слуховой и моторной коры, а также внутреннего коленчатого тела у кроликов.
В опытах на зрительной коре морской свинки было показано, что тормозные влияния осуществляются от периферических частей микроансамблей нейронов к их центральным частям.
По поводу нейронов, тормозящихся даже при осуществлении условных рефлексов, было высказано предположение, что они представляют особую форму внутреннего торможения, которая сопутствует каждому акту условно-рефлекторной деятельности, и обеспечивают его приспособительную точность. Однако такое определение роли тормозящихся нейронов встретило возражение, согласно которому в описываемой ситуации речь идет не о внутреннем, а о координационном торможении, обеспечивающем точность выполнения условного рефлекса.
Таким образом,
Нейрофизиологический механизм внутреннего торможения. О нейрофизиологии внутреннего торможения высказывались разные предположения. Так, И.П. Павлов (1927), не решая окончательно вопрос о его природе, считал возможным рассматривать развитие внутреннего торможения при неподкреплении как результат перевозбуждения клеток коркового представительства сигнала, которые при подкреплении предохранялись от этого затормаживанием. Исследование свойств угасательного торможения привело к сходному заключению о том, что оно является фазой перевозбуждения возбудительного очага. Другое предположение исходило из того, что при неподкреплении ослабляются прямые временные связи от зоны корковой проекции условного раздражителя к зоне безусловного и усиливаются обратные, по которым активируются апикальные дендриты выходных пирамидных нейронов в зоне представительства безусловного рефлекса. Такое локальное возбуждение дендритов создает электротоническую блокаду синапсов, и рефлекс прекращается (И.С. Беритов, 1956). Другим источником локального возбуждения апикальных дендритов пирамид могут быть неспецифические структуры таламуса (А.И. Ройтбак, 1958).
Исходя из того, что рабочее состояние высших отделов мозга обусловливается тоническим восходящим активирующим влиянием ретикулярной формации среднего мозга, условное торможение рассматривали как результат снижения этого влияния. Дальнейшие исследования показали, что для развития внутреннего торможения. Важное значение имеет изменение тонуса ретикулярной формации (Б.И. Котляр, 1977). Однако это не единственная причина внутреннего торможения.
По другому предположению, ведущую роль в угнетении условных рефлексов играет антагонист мезэнцефалической системы — восходящая «синхронизирующая» таламокортикальная система (Г. Мегун, 1956). Считают также, что поскольку автостабилизация корковых процессов поддерживается «превентивным» торможением (П.В. Симонов, 1965), его углубление при действии неподкрепляемых раздражений образует внутреннее торможение.
На основании сходства ряда электрофизиологических показателей реализации положительных и отрицательных условных рефлексов, особенно при тонких дифференцировках, условное торможение рассматривают как возбуждение, ослабленное отсутствием обратной афферентации от исполнительных органов ввиду отсутствия подкрепления (Ю.Г. Кратин, 1967). Неподкрепление условного рефлекса само по себе становится сигналом «неблагополучия» и создает «трудное состояние», возникновение которого по механизму внешнего торможения задерживает рефлекс (П.К. Анохин, 1968). Различия порогов реакций, вызываемых безусловным и условным раздражителем, объясняют частичным торможением центральных структур собственного рефлекса на раздражитель, который сделали условным, а отмена подкрепления нарушает этот механизм, вызывает перевозбуждение и охранительное торможение в элементах временной связи (Э.А. Асратян, 1970).
Существует мнение, что внутреннее торможение есть не что иное, как образование нового рефлекса, в котором подкреплением является отсутствие безусловного раздражителя (Ю. Конорски, 1970). Условное торможение по многим своим проявлениям и электрофизиологическим показателям обнаруживает свойства доминанты, которая становится очагом нераспространяющихся возбуждений (В.С. Русинов, 1969). Если образование условного рефлекса происходит путем «прокладывания связей» между сигналом и доминантным очагом безусловного рефлекса, то при отсутствии подкрепления предполагают накопление в представительстве сигнального стимула нераспространяющегося возбуждения, образующего своеобразный локальный доминантный очаг. Поскольку образование пути условного возбуждения определяется пространственной синхронизацией и когерентностью ритмов активности (М.Н. Ливанов, 1972), их рассогласование препятствует распространению возбуждения по путям временных связей. Этому соответствуют данные о том, что при выработке дифференцировки двигательного рефлекса на звук коэффициенты корреляции импульсных потоков нейронов слуховой и ассоциативной коры кошки ранее положительные становятся отрицательными.
На основании результатов опытов, в которых изучали влияние симпатической части нервной системы на высшую нервную деятельность, был сделан вывод о ее участии в механизме формирования внутреннего торможения. Было высказано предположение, что выработка дифференцировочного торможения представляет собой образование нового рефлекса с подкреплением из памяти. Исследование процессов внутреннего торможения на нейронном и системном уровнях привело к заключению о существенной роли в его механизме явлений возвратного торможения и развития гиперполяризационных процессов в коре. Недавно представлены факты, свидетельствующие о том, что при изучении механизма внутреннего торможения следует учитывать изменения тонуса активности нейронов коры и подкорковых структур головного мозга.
Если относить к внутреннему все виды торможения, которые вырабатываются, то к нему принадлежит и торможение, возникающее при многократном повторении любого афферентного раздражения, которое приводит к угашению ориентировочной реакции на него. Такие явления «привыкания», или «габитуации», в сущности представляют собой случаи угашения натурального условного рефлекса. За последнее время они детально изучены на нейронном и молекулярном уровнях на гигантских нейронах беспозвоночных. При этом найдено, что угашение реакции втягивания жабер у аплизии сопровождается снижением моносинаптических ВПСП мотонейрона в результате уменьшения выброса медиатора из пресинаптических терминалей в синаптическую щель. Исследование квантового состава выделяющегося в синапс медиатора на примерах привыкания нервных клеток виноградной улитки и нейронов коры и гиппокампа кролика показало, что во всех случаях имело место уменьшение числа, а в некоторых случаях и объема квантов.
Несмотря на обилие работ по изучению внутреннего торможения, остается еще много неясного в его механизмах.
| © 2025 Библиотека RealLib.org (support [a t] reallib.org) |