"Основы физиологии высшей нервной деятельности" - читать интересную книгу автора (Коган Александр Борисович)
Глава 2 ОБРАЗОВАНИЕ УСЛОВНЫХ РЕФЛЕКСОВ
Основной элементарный акт высшей нервной деятельности — образование условного рефлекса. Здесь эти свойства будут рассмотрены, как и все общие законы физиологии высшей нервной деятельности, на примере условных слюноотделительных рефлексов собаки.
Условный рефлекс занимает высокое место в эволюции временных связей, являющихся универсальным приспособительным явлением в животном мире. Наиболее примитивный механизм индивидуального приспособления к меняющимся условиям жизни, видимо, представляют
Существует бесчисленное множество условных рефлексов. При соблюдении соответствующих правил любой воспринимаемый раздражитель можно сделать стимулом, запускающим условный рефлекс (сигналом), а любую деятельность организма — его основой (подкреплением). По роду сигналов и подкреплений, а также по отношениям между ними созданы разные классификации условных рефлексов. Что касается изучения физиологического механизма временных связей, то здесь исследователям предстоит большая работа.
На примере систематического изучения слюноотделения у собак наметились общие признаки условного рефлекса, а также частные признаки разных категорий условных рефлексов. Определилась классификация условных рефлексов по следующим частным признакам: 1) обстоятельства образования, 2) род сигнала, 3) состав сигнала, 4) род подкрепления, 5) отношение во времени условного раздражителя и подкрепления.
Общие признаки условных рефлексов. Какие признаки являются общими и обязательными для всех условных рефлексов? Условный рефлекс а) является индивидуальным высшим приспособлением к меняющимся условиям жизни; б) осуществляется высшими отделами центральной нервной системы; в) приобретается путем вре́менных нервных связей и утрачивается, если вызвавшие его условия среды изменились; г) представляет собой предупредительную сигнальную реакцию.
Итак,
Натуральные и искусственные условные рефлексы. В зависимости от характера сигнального раздражителя условные рефлексы делят на натуральные и искусственные.
Примером натурального условного пищевого рефлекса может служить выделение слюны у собаки на запах мяса. Этот рефлекс с неизбежностью образуется естественным путем в течение жизни собаки.
Натуральные условные рефлексы природа вырабатывает из поколения в поколение у всех животных соответственно их образу жизни. В результате
Избирательная чувствительность к экологически адекватным сигналам проявляется в реакциях нервных клеток мозга.
Экстероцептивные, интероцептивные и проприоцептивные условные рефлексы. Условные рефлексы на внешние раздражители называют
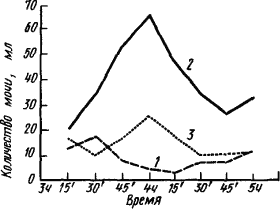 |
Рис. 1. Интероцептивный условный рефлекс мочеобразования при «мнимом вливании» физиологического раствора (по К. Быкову):
1 — исходная кривая мочеобразования, 2 — мочеобразование в результате вливания в желудок 200 мл физиологического раствора, 3 — мочеобразование в результате «мнимого вливания» после 25 истинных
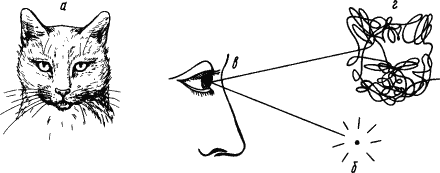 |
Рис. 2. Изучение проприоцептивных компонентов зрительного представления человека:
Особую категорию условных рефлексов составляют модельные опыты с электрическим раздражением мозга в качестве подкрепления или сигнала; с использованием в качестве подкрепления ионизирующей радиации; созданием доминанты; выработкой временных связей между пунктами нейронально-изолированной коры; исследованием суммационного рефлекса, а также образованием условных реакций нервной клетки на сигнал, подкрепляемый локальной электрофоретической аппликацией медиаторов.
Условные рефлексы на простые и комплексные раздражители. Как было показано, условный рефлекс можно выработать на какой-либо один из перечисленных экстеро-, интеро- или проприоцептивных раздражителей, например на включение света или на простой звук. Но в жизни это бывает редко. Чаще сигналом становится комплекс из нескольких раздражителей, например запах, тепло, мягкая шерсть матери-кошки становятся раздражителем условного сосательного рефлекса для котенка. Соответственно разделяют условные рефлексы на
Условные рефлексы на простые раздражители не требуют пояснений. Условные рефлексы на сложные раздражители делятся по признаку отношений между членами комплекса (рис. 3).
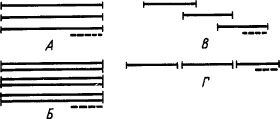 |
Рис. 3. Отношение во времени между членами комплексов сложных условных раздражителей.
одинарными линиями показаны индифферентные раздражители, двойными — ранее выработанные сигналы, пунктиром — подкрепление
Условные рефлексы, выработанные на основе различных подкреплений. Основой для образования условного рефлекса — его
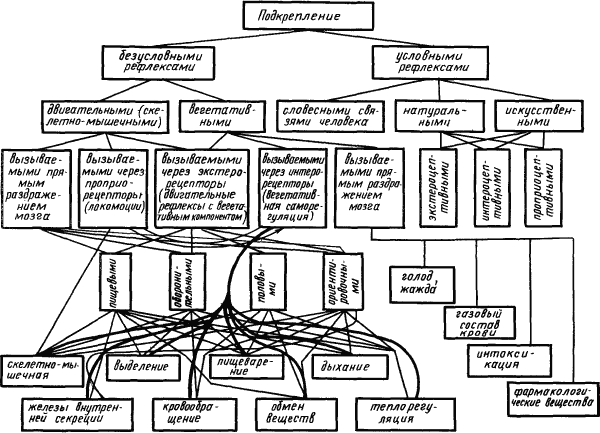 |
Рис. 4. Классификация подкреплений, на которые могут образовываться условные рефлексы
Каждый условный рефлекс, в свою очередь, может стать основой для образования нового условного рефлекса. Новая условная реакция, выработанная с помощью подкрепления сигнала другим условным рефлексом, называется
Условные рефлексы второго, третьего и далее порядков широко распространены в природе. Они составляют наиболее значительную и совершенную часть натуральных условных рефлексов. Например, когда волчица кормит волчонка мясом растерзанной добычи, у него вырабатывается натуральный условный рефлекс первого порядка. Вид и запах мяса становятся для него сигналом еды. Затем он «учится» охоте. Теперь эти сигналы — вид и запах мяса пойманной добычи — играют роль основы для выработки охотничьих приемов подстерегания и преследования живой добычи. Так приобретают свое вторичное сигнальное значение разнообразные охотничьи признаки: обглоданный зайцем кустик, следы отбившейся от стада овцы и т.д. Они становятся раздражителями условных рефлексов второго порядка, выработанных на основе натуральных.
Наконец, исключительное разнообразие условных рефлексов, подкреплением которых служат другие условные рефлексы, встречается в высшей нервной деятельности человека. Более подробно они будут рассмотрены в гл. 17. Здесь лишь необходимо отметить, что в отличие от условных рефлексов животных
Условные рефлексы, выработанные при разном соответствии во времени сигнала и подкрепления. По тому, как располагается во времени сигнал относительно подкрепляющей реакции, различают
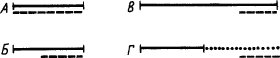 |
Рис. 5. Варианты временного соотношения сигнала и подкрепления.
сплошной линией обозначено время действия сигнала, прерывистой линией — время подкрепления
Какие условия должны быть соблюдены, чтобы деятельность высших отделов центральной нервной системы могла завершаться выработкой условного рефлекса?
Сочетание сигнального раздражителя с подкреплением. Это условие выработки временных связей выявилось с первых же опытов со слюнными условными рефлексами. Шаги служителя, несущего пищу, только тогда вызывали «психическое слюноотделение», когда они сочетались с едой.
Этому не противоречит образование следовых условных рефлексов. Подкрепление сочетается в таком случае со следом возбуждения нервных клеток от ранее включенного и уже выключенного сигнала. Но если подкрепление станет опережать индифферентный раздражитель, то условный рефлекс удается выработать с большим трудом, лишь принимая ряд специальных мер. Это и понятно, так как если собаку сначала покормить, а потом дать сигнал еды, то, строго говоря, он даже не может называться сигналом, так как не предупреждает о предстоящих событиях, а отражает прошедшее. При этом безусловный рефлекс подавляет сигнальное возбуждение и препятствует образованию условного рефлекса на такой раздражитель.
Индифферентность сигнального раздражителя. Агент, выбранный в качестве условного раздражителя пищевого рефлекса, не должен сам по себе иметь какое-либо отношение к пище. Он должен быть безразличным, т.е. индифферентным, для слюнных желез. Сигнальный раздражитель не должен вызывать значительной ориентировочной реакции, мешающей образованию условного рефлекса. Однако ориентировочную реакцию вызывает каждый новый раздражитель. Поэтому для утраты им новизны его необходимо применять повторно. Лишь после того, как ориентировочная реакция будет практически погашена или сведена к незначительной величине, начинается образование условного рефлекса.
Преобладание силы возбуждения, вызванного подкреплением. Сочетание стука метронома и кормления собаки ведет к быстрому и легкому образованию у нее условного слюноотделительного рефлекса на этот звук. Но если пытаться сочетать с едой оглушительный стук механической трещотки, то такой рефлекс образовать чрезвычайно трудно. Для выработки временной связи большое значение имеет соотношение силы сигнала и подкрепляющей реакции. Чтобы между ними образовалась временная связь, очаг возбуждения, создаваемый последней, должен быть сильнее очага возбуждения, создаваемого условным раздражителем, т.е. должна возникать доминанта. Лишь тогда произойдет распространение возбуждения от очага индифферентного раздражителя к очагу возбуждения от подкрепляющего рефлекса.
Необходимость значительной интенсивности возбуждения именно подкрепляющей реакции имеет глубокий биологический смысл. В самом деле, условный рефлекс — это предупредительная реакция на сигнал о предстоящих существенных событиях. Но если раздражитель, который хотят сделать сигналом, оказывается событием даже более существенным, чем те, которые за ним последуют, то этот раздражитель сам вызывает соответствующую реакцию организма.
Отсутствие посторонних раздражителей. Каждое постороннее раздражение, например неожиданно донесшийся шум, вызывает уже упоминавшуюся ориентировочную реакцию. Собака настораживается, поворачивается в направлении звука и, главное, прекращает свою текущую деятельность. Животное все обращено навстречу новому раздражителю. Недаром И.П. Павлов называл ориентировочную реакцию рефлексом «Что такое?». Напрасно в это время экспериментатор будет подавать сигнал и предлагать собаке пищу. Условный рефлекс будет задержан более важным в данный момент для животного — ориентировочным рефлексом. Эта задержка создается дополнительным очагом возбуждения в коре мозга, который тормозит условное возбуждение и препятствует образованию временной связи. В природе множество таких случайностей влияет на ход образования условных рефлексов животных. Отвлекающая обстановка снижает продуктивность и умственной работы человека.
Нормальная работоспособность нервной системы. Полноценная замыкательная функция возможна при условии, если высшие отделы нервной системы находятся в нормальном рабочем состоянии. Метод хронического эксперимента потому и дал возможность обнаружить и изучить процессы высшей нервной деятельности, что при этом сохранялось нормальное состояние животного. Работоспособность нервных клеток мозга резко снижается при недостаточном питании, при действии ядовитых веществ, например бактериальных токсинов при болезнях, и т.д. Поэтому общее состояние здоровья является важным условием нормальной деятельности высших отделов мозга. Всем известно, как это условие влияет на умственную работу человека.
На образование условных рефлексов существенное влияние оказывает состояние организма. Так, физическая и умственная работа, условия питания, активность гормонов, действие фармакологических веществ, дыхание при повышенном или пониженном давлении, механические перегрузки и ионизирующее излучение в зависимости от интенсивности и сроков воздействия могут видоизменить, усилить или ослабить условно-рефлекторную деятельность вплоть до полного ее подавления.
Образование условных рефлексов и реализация актов высшей нервной деятельности в чрезвычайной степени зависят от потребности организма в биологически значимых агентах, используемых в качестве подкрепления. Так, у сытой собаки очень трудно выработать пищевой условный рефлекс, она будет отворачиваться от предлагаемой пищи, а у голодного животного с высокой пищевой возбудимостью он образуется быстро. Общеизвестно, как интерес учащегося к предмету занятий способствует лучшему его усвоению. Эти примеры показывают большое значение фактора отношения организма к проявляемым раздражителям, который обозначается как
Исследование конечных, поведенческих проявлений высшей нервной деятельности значительно опередило изучение ее внутренних механизмов. До настоящего времени еще недостаточно изучены как структурные основы временной связи, так и ее физиологическая природа. По этому поводу высказываются разные взгляды, но вопрос еще не решен. Для его решения ведутся многочисленные исследования на системном и клеточном уровнях; используют электрофизиологические и биохимические показатели динамики функционального состояния нервных и глиальных клеток с учетом результатов раздражения или выключения различных структур мозга; привлекают данные клинических наблюдений. Однако на современном уровне исследований становится все более определенным, что при этом наряду со структурной необходимо учитывать и нейрохимическую организацию мозга.
Изменение локализации замыкания временных связей в эволюции. Независимо от того, считать ли, что условные реакции
Уже на раннем этапе эволюции позвоночных в передних отделах первоначально однородной мозговой трубки выделяется головной мозг, управляющий приспособительным поведением. В нем развиваются структуры, имеющие наибольшее значение для замыкания вредных связей в процессе условно-рефлекторной деятельности. На сновании опытов с удалением частей мозга у
У
У
Удаление частей и всей коры больших полушарий. Если у взрослой собаки удалить затылочные области коры, то она утрачивает все сложные зрительные условные рефлексы и не может их восстановить. Такая собака не узнает своего хозяина, безразлична к виду самых лакомых кусков пищи, равнодушно смотрит на пробегающую мимо кошку, которую прежде бросилась бы преследовать. Наступает то, что раньше называли «психической слепотой». Собака видит, так как она обходит препятствия, поворачивается к свету. Но она «не понимает» смысла увиденного. Без участия зрительной коры зрительные сигналы остаются ни с чем не связанными.
И все-таки такая собака может образовывать очень простые зрительные условные рефлексы. Например, появление освещенной фигуры человека можно сделать сигналом пищи, вызывающим выделение слюны, облизывание, виляние хвостом. Следовательно, и в других областях коры находятся клетки, воспринимающие зрительные сигналы и способные их связывать с теми или иными действиями. Эти факты, подтвержденные и в опытах с повреждением корковых областей представительства других сенсорных систем, привели к мнению, что проекционные зоны перекрывают друг друга (Л. Лючиани, 1900). Дальнейшие исследования вопроса о локализации функций в коре в работах И.П. Павлова (1907–1909) показали широкое перекрытие проекционных зон, зависящее от природы сигналов и образуемых временных связей. Обобщая все эти исследования, И.П. Павлов (1927) выдвинул и обосновал представление о
Рассеянные элементы не в состоянии заменить специализированные клетки ядра для образования тонких временных связей. Собака после удаления затылочных долей может вырабатывать лишь самые простые условные рефлексы, например, на вид освещенной фигуры. Заставить же ее различать две такие фигуры, сходные по форме, не удается. Однако если удаление затылочных долей произвести в раннем возрасте, когда еще не произошло выделение и закрепление проекционных зон, то, вырастая, эти животные обнаруживают способность к выработке сложных форм условных зрительных рефлексов.
Возможность широкой взаимозаменяемести функций коры мозга в раннем онтогенезе соответствует свойствам малодифференцированной коры мозга млекопитающих в филогенезе. С этой точки зрения получают объяснение результаты опытов на крысах, у которых степень нарушения условных рефлексов оказалась зависящей не от конкретной области удаляемой коры, а от общего объема удаляемой корковой массы (рис. 6). На основании этих опытов было сделано заключение, что для условно-рефлекторной деятельности все отделы коры имеют одинаковое значение, кора
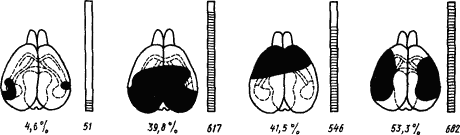 |
Рис. 6. Взаимозаменяемость частей коры мозга после их удаления у крыс (по К. Лешли):
удаленные области зачернены, цифрами под мозгом обозначена величина удаления в процентах ко всей поверхности коры, цифрами под столбиками — число ошибок при испытании в лабиринте
Первые опыты с удалением всей коры больших полушарий (lt;…пропуск…gt;Гольтц, 1982) показали, что после такой обширной операции, по-видимому, затрагивающей и ближайшую подкорку, собаки ничему не могли научиться. В опытах на собаках с удалением коры без травмирования подкорковых структур мозга удалось выработать
Разработка операции перерезки всех восходящих и нисходящих путей, связывающих кору с другими образованиями мозга, позволила осуществить декортикацию без прямого травмирования подкорковых структур и изучить роль коры в условно-рефлекторной деятельности. Оказалось, что у этих кошек можно было с большим трудом вырабатывать только грубые условные рефлексы общих движений, а оборонительного условного сгибания лапы не удалось получить даже после 150 сочетаний. Однако уже после 20 сочетаний на сигнал появилась реакция изменения дыхания и некоторые условные вегетативные реакции.
Конечно, при всех хирургических операциях трудно исключить их травмирующее влияние на подкорковые структуры и быть уверенным, что утраченная способность к тонкой условно-рефлекторной деятельности была функцией коры. Убедительные доказательства дали опыты с временным обратимым выключением функций коры, которое проявляется в распространяющейся депрессии электрической активности при аппликации KCI на ее поверхность. При выключении таким образом коры мозга крысы и испытании в это время реакции животного на условные и безусловные раздражители можно видеть, что безусловные рефлексы полностью сохраняются, а условные — нарушаются. Как видно из рис. 7, более сложные оборонительные и особенно пищевые условные рефлексы при максимальной депрессии в течение первого часа полностью отсутствуют, а простая защитная реакция избегания страдает в меньшей степени.
Таким образом, результаты экспериментов с частичной и полной хирургической и функциональной декортикацией говорят о том, что у
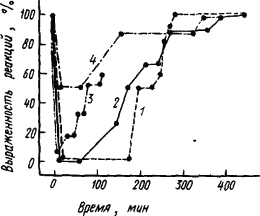 |
Рис. 7. Влияние временного выключения коры путем распространяющейся депрессии на пищевые
Корково-подкорковые отношения в процессах высшей нервной деятельности. Современные исследования подтверждают высказывание И.П. Павлова, что условно-рефлекторная деятельность осуществляется совместной работой коры и подкорковых структур. Из рассмотрения эволюции мозга как органа высшей нервной деятельности следует, что способность к образованию временных связей, обеспечивающих приспособительное поведение, проявляли структуры промежуточного мозга у рыб и стриарных (полосатых) тел у птиц, представляющие собой филогенетически наиболее молодые его отделы. Когда же у млекопитающих над этими отделами мозга возникла филогенетически самая молодая новая кора, осуществляющая наиболее тонкий анализ сигналов, то к ней перешла ведущая роль в образовании временных связей, организующих приспособительное поведение.
Структуры мозга, оказавшиеся подкорковыми, сохраняют в какой-то мере свои способности к замыканию временных связей, которые обеспечивают приспособительное поведение, свойственное тому уровню эволюции, когда эти структуры были ведущими. Об этом свидетельствует описанное выше поведение животных, которые после выключения коры мозга могли с трудом вырабатывать лишь очень примитивные условные рефлексы. Вместе с тем возможно, что такие примитивные временные связи не потеряли полностью своего значения и образуют часть нижнего уровня сложного иерархического механизма высшей нервной деятельности, возглавляемого корой больших полушарий.
Взаимодействие коры и подкорковых отделов мозга осуществляется и путем
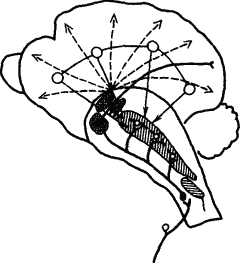 |
Рис. 8. Взаимодействие ретикулярной формации среднего мозга и коры (по Л.Г. Воронину):
жирными линиями обозначены афферентные специфические пути с коллатералями к ретикулярной формации, прерывистыми — восходящие пути к коре, тонкими — влияние коры на ретикулярную формацию, вертикальной штриховкой — облегчающая зона, горизонтальной — тормозная зона, клеточной штриховкой — таламические ядра
Иное действие на функциональное состояние коры оказывают
Результаты некоторых экспериментов истолковывали как доказательство тормозящего действия стриарных структур на поведение животных. Однако дальнейшие исследования, в частности опыты с разрушением и стимуляцией хвостатых тел, и другие факты привели к заключению о наличии более сложных корково-подкорковых отношений.
Некоторые исследователи рассматривают факты об участии подкорковых структур в процессах высшей нервной деятельности как основание для того, чтобы считать их местом замыкания временных связей. Так возникло представление о
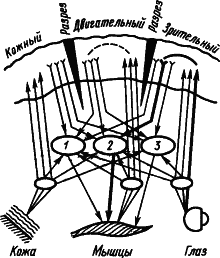 |
Рис. 9. Множественное замыкание временных связей в коре (показано пунктиром), которым не препятствуют ее разрезы (по А.Б. Когану):
Как показали многочисленные исследования, участие подкорковых структур в процессах высшей нервной деятельности не ограничивается регуляторной ролью ретикулярной формации среднего мозга и лимбических структур. Ведь уже на подкорковом уровне происходят анализ и синтез действующих раздражителей и оценка их биологического значения, в значительной мере определяющая характер образующихся связей с сигналом. Использование показателей формирования наиболее коротких путей, по которым сигнал достигает разных подкорковых структур мозга, выявило наиболее выраженное участие в процессах обучения задних отделов таламуса и поля CA3 гиппокампа. Роль гиппокампа в явлениях памяти подтверждается многими фактами. Наконец, нет оснований предполагать, что способность к примитивной замыкательной деятельности структур мозга, которая приобреталась в эволюции, когда они были ведущими, теперь у них полностью исчезла, когда эта функция перешла к новой коре.
Таким образом, корково-подкорковые отношения определяются
Межполушарные отношения. Каким образом полушария головного мозга, являющиеся парным органом, участвуют в процессах образования условных связей? Ответ на этот вопрос был получен в опытах на животных, которым была сделана операция «расщепления» мозга путем перерезки мозолистого тела и передней комиссуры, а также продольного разделения перекреста зрительных нервов (рис. 10). После такой операции можно было вырабатывать разные условные рефлексы правого и левого полушарий, показывая разные фигуры правому или левому глазу. Если у оперированной таким образом обезьяны выработать условный рефлекс на световой стимул, подаваемый на один глаз, а затем подать его на другой глаз, то никакой реакции не последует. «Обучение» одного полушария оставляло другое «необученным». Однако при сохранении мозолистого тела и другое полушарие оказывается «обученным». Мозолистое тело осуществляет
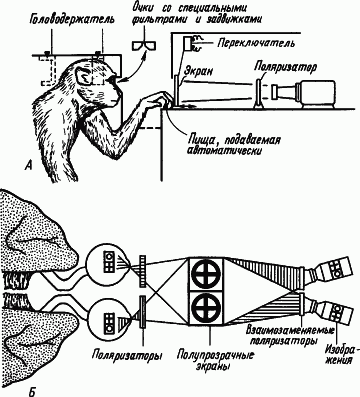 |
Рис. 10. Исследования процессов обучения у обезьян, подвергнутых операции «расщепления» мозга.
С помощью метода функционального выключения коры мозга у крыс воспроизводили на некоторое время условия «расщепленного» мозга. При этом временные связи могло образовать одно оставшееся активным полушарие. Этот рефлекс проявлялся и после прекращения действия распространяющейся депрессии. Он сохранился даже после инактивации полушария, которое было активным во время выработки этого рефлекса. Следовательно, «обученное» полушарие передало по волокнам мозолистого тела приобретенный навык «необученному». Однако этот рефлекс исчезал, если такую инактивацию производили до того, как полностью восстановится активность полушария, включенного во время выработки условного рефлекса. Таким образом, для переноса приобретенного навыка из одного полушария в другое необходимо, чтобы оба они были активны.
Дальнейшие исследования межполушарных отношений при образовании временных связей условных рефлексов показали, что во взаимодействии полушарий специфическую роль играют процессы торможения. Так, полушарие, противоположное стороне подкрепления, становится доминирующим. Оно вначале осуществляет формирование приобретенного навыка и его перенос в другое полушарие, а потом, притормаживая активность противоположного полушария и оказывая избирательно тормозное влияние на структуру временных связей, совершенствует условный рефлекс.
Таким образом,
Предположения о месте замыкания временных связей в больших полушариях головного мозга. Открыв условный рефлекс, И.П. Павлов сначала предположил, что временная связь является «вертикальной связью» между зрительной, слуховой или иными частями коры больших полушарий и подкорковыми центрами безусловных рефлексов, например пищевого —
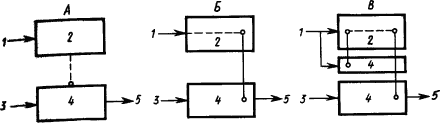
Наличие в коре больших полушарий собаки представительства безусловных рефлексов доказывается следующими фактами. Если применять в качестве пищевого раздражителя сахар, то слюноотделение на него вырабатывается лишь постепенно. Если какой-либо условный раздражитель не подкреплять, то следующее за ним «сахарное» слюноотделение уменьшается. Значит, этот безусловный рефлекс имеет нервные клетки, расположенные в сфере протекания корковых процессов. Дальнейшие исследования показали, что если удалить у собаки кору, то ее безусловные рефлексы (слюнные, отделение желудочного сока, движения конечностей) претерпевают стойкие изменения. Следовательно, безусловные рефлексы кроме подкоркового центра имеют центры и на корковом уровне. Вместе с тем раздражитель, который сделан условным, также имеет представительство в коре. Отсюда возникло предположение (Э.А. Асратян, 1963), что временные связи условного рефлекса замыкаются между этими представительствами (рис. 11,
 |
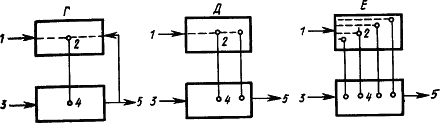 |
Рис. 11. Различные предположения о структуре временной связи условного рефлекса (объяснение см. в тексте):
Рассмотрение процессов замыкания временных связей как центральных звеньев формирования функциональной системы (П.К. Анохин, 1961) относит замыкание к структурам коры, где происходит сопоставление содержания сигнала —
Изучение двигательных условных рефлексов показало сложную структуру образующихся при этом временных связей (Л.Г. Воронин, 1952). Каждое совершающееся по сигналу движение само становится сигнальным для складывающихся при этом двигательных координаций. Образуются две системы временных связей: на сигнал и на движение (рис. 11,
Наконец, исходя из того, что условные рефлексы сохраняются при хирургическом разделении сенсорных и моторных корковых областей и даже после множественных разрезов коры, а также учитывая, что кора обильно снабжена как входящими, так и выходящими путями, было высказано предположение, что замыкание временных связей может происходить в каждом ее микроучастке между его афферентными и эфферентными элементами, которые активируют центры соответствующих безусловных рефлексов, служащих подкреплением (А.Б. Коган, 1961) (см. рис. 9 и 11,
Нейронная структура временной связи в коре больших полушарий. Современные сведения о микроскопическом строении коры больших полушарий в сочетании с результатами электрофизиологических исследований позволяют с известной степенью вероятности судить о возможном участии тех или иных корковых нейронов в образовании временных связей.
Высокоразвитая кора мозга млекопитающих, как известно, делится на шесть слоев разного клеточного состава. Приходящие сюда нервные волокна заканчиваются большей частью у клеток двух видов. Один из них — это вставочные нейроны, расположенные во
Другой вид клеток, с которыми приходящие волокна образуют наибольшее число контактов, составляют кустовидно ветвящиеся округлые и угловатые короткоотростчатые клетки, часто имеющие звездчатую форму. Они располагаются главным образом в
Система вставочных и звездчатых нейронов может вступать в бесчисленное множество контактов с ассоциативными и проекционными крупными нейронами пирамидной формы, расположенными в
Согласно такому представлению о функциях корковых клеток замыкание временной связи осуществляется в системе звездчатых, вставочных и ассоциативных нейронов. Возможны два типа замыкания. Один из них — это замыкание с участием преимущественно зрительных, слуховых и других
Расположение различных корковых нейронов на путях прохождения импульсов и динамика функциональных показателей их активности при образовании условного рефлекса позволяет предположить, что замыкание происходит главным образом на клеточных телах вставочных нейронов. Возможно, особая роль при этом принадлежит нейронам с распространенными рецептивными полями. Существует мнение, что в онтогенезе процессы замыкания зависят от созревания множественных выростов на дендритах нейронов —
Известны многочисленные попытки выделить по свойствам нейронов клетки, участвующие в образовании временных связей. Такими свойствами считали высокую частоту фоновой импульсации, большую реакцию на условный раздражитель, способность реагировать на раздражители разной модальности, полисенсорность, высокую лабильность, способность к пластическим перестройкам — избирательную полифункциональность, сходство реакции на условный и безусловный раздражители, ответы преимущественно на подкрепление. На основании опытов главным образом с нервной системой моллюсков возникло представление о ведущей роли в образовании новых форм поведения так называемых
Что же происходит с нейронами, участвующими в замыкании временной связи? Давно замечено, что после выработки условного рефлекса возрастает и величина безусловного рефлекса, хотя сила подкрепляющего раздражителя остается прежней. Отсюда следует, что выработка условного рефлекса связана с повышением возбудимости нервных клеток, образующих новый рефлекторный путь. Повышение возбудимости корковых структур, участвующих в образовании условно-рефлекторной связи, можно обнаружить и непосредственно, измеряя пороги их раздражения.
После вживления электродов в область двигательного анализатора (например, кошки), можно измерять порог прямого раздражения и определять изменения возбудимости исследуемых нервных клеток мозга у животного при выработке условного рефлекса. Оказалось, что процесс выработки двигательного условного рефлекса выражается прежде всего стойким повышением возбудимости клеток двигательного анализатора (рис. 12). Только когда возбудимость корковых клеток повышается до известного уровня, появляются двигательные признаки условного рефлекса — сгибание лапы. Каждый очередной сигнал вызывает быстрый дополнительный подъем возбудимости нейронов двигательного анализатора. Если же отменить условный рефлекс, то возбудимость нервных клеток снижается до уровня, предшествующего началу выработки рефлекса. Соответственно изменяются импульсация и ответы на условный раздражитель. При этом меняется деятельность нейронов.
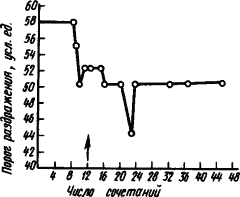 |
Рис. 12. Изменение порога прямого раздражения сенсомоторной коры при выработке условного рефлекса сгибания лапы у кошки (по Н.И. Николаевой):
стрелкой обозначен момент образования условного рефлекса
Так, полисенсорные нейроны, которые наиболее часто перестраиваются при образовании условного рефлекса, могут стать моносенсорными или не участвовать в условных реакциях. Исходя из того что при разных видах условно-рефлекторной деятельности в разрядах нейронов лобной коры доминируют межимпульсные интервалы каждый раз определенной длительности, было высказано предположение что положительный условный рефлекс и его торможение кодируются различными классами межимпульсных интервалов.
Электрические потенциалы, возникающие при деятельности нервной системы, могут служить тонким показателем происходящих в ней процессов. Поэтому в изучении процессов высшей нервной деятельности электрофизиологические исследования занимают особое место. Такие исследования проводят с использованием показателей фоновой ритмики (активности) и вызванных потенциалов разных структур мозга, импульсных разрядов и мембранных потенциалов отдельных нервных клеток. Сопоставление полученных данных в сочетании с показателями условных рефлексов позволяет выявить некоторые сведения о механизмах, свойствах образования и осуществления временных связей.
Фоновая ритмика (активность). Непрерывно идущие колебания потенциалов мозга, обозначаемые как фоновая (спонтанная) или основная ритмика, меняют свой характер, отражая течение процессов высшей нервной деятельности. Это явление было обнаружено случайно. После того как несколько раз фотографировали с экрана осциллографа
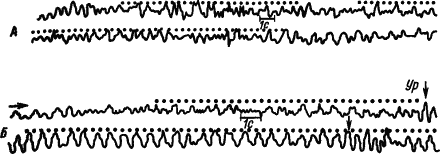 |
Рис. 13. Синхронизация потенциалов моторной коры мозга кролика ритмическими вспышками света, совпадающими по частоте с электрокожным раздражением лапы при выработке двигательного оборонительного рефлекса.
в каждой паре записей нижняя продолжает верхнюю, точками обозначены вспышки света, Ур — условно-рефлекторное движение лапы
Дальнейшие исследования показали, что реакция десинхронизации электрокортикограммы (ЭКоГ) человека и животных, так же как реакция усвоения ритма, может отражать многие свойства нервных механизмов условно-рефлекторной деятельности. Так, во время генерализации условных рефлексов эти реакции возникают на обширной территории коры, охватывая области проекций и условного и безусловного раздражителей, а по мере специализации условных рефлексов сосредоточиваются на более ограниченной зоне, большей частью в двигательной области коры.
В процессе образования временной связи реакции синхронизации и усвоения ритмов раздражителя вначале формируются в корковых проекциях сигнала, затем в зоне проекции подкрепления, при дальнейшем упрочении условного рефлекса идут на убыль в зоне проекции сигнала, но долго сохраняются в зоне проекции подкрепления. При этом усвоение ритма происходит сначала в верхних, а потом в нижних слоях коры. Парная работа полушарий в формировании временных связей проявляется в том, что по мере выработки условного рефлекса ритмы сигнального стимула усваиваются вначале структурами верхних слоев коры полушария, контралатерального стороне подкрепления, затем они появляются в его нижних слоях и во всех слоях ипсилатерального полушария, потом они уменьшаются и исчезают всюду, кроме нижних слоев полушария, контралатерального стороне подкрепления (рис. 14).
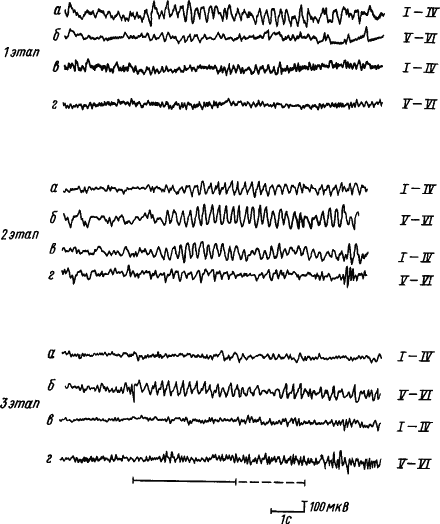 |
Рис. 14. Пространственная динамика усвоения ритма условного раздражителя в слоях симметричных пунктов коры больших полушарий головного мозга в процессе выработки условного рефлекса (по А.Б. Когану, Г.А. Кураеву):
Разработка методов электроэнцефалографии с одновременной регистрацией ЭКоГ от десятков и сотен пунктов коры и использование электронно-вычислительных машин позволили выяснить роль синхронизации ритмов биоэлектрической активности в областях корковой проекции условного и безусловного раздражителей для образования временной связи между ними. Показано, например, что условные оборонительные движения лапы у кролика проявлялись тогда, когда коэффициент корреляции ритмов активности исследуемых корковых нейронов достигал достаточно высокого значения. Это подтверждено и для спонтанных движений. Функциональное значение пространственной синхронизации электрических потенциалов головного мозга было показано на многочисленных примерах. Показатели синхронизации также демонстрируют иррадиацию нервных процессов при начальной генерализации условного рефлекса, а по мере его закрепления и специализации нарастающую их концентрацию (рис. 15).
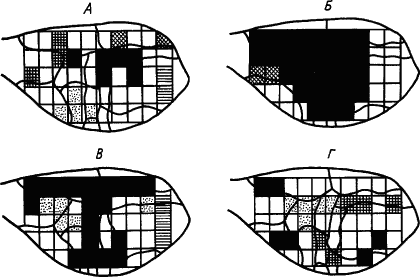 |
Рис. 15. Динамика синхронизации волн электрокортикограммы кролика в процессе выработки условного рефлекса.
квадратами обозначены участки отведения; одинаково изображены участки, синхронность волн между которыми сохраняется не менее 70 % времени измерения
Многие сведения, существенные для понимания механизма образования временных связей, были получены при исследовании разных режимов условного раздражения и подкрепления, а также пространственных характеристик регистрируемой активности. Так, успех образования временной связи по показателям синхронизации усвоенных ритмов оказался зависимым главным образом от четырех факторов: 1) изоритмии сигнала и подкрепления; 2) оптимальности их частоты (для кролика 4–7 Гц); 3) отношения фаз (синфазность) и 4) достаточного объема нервного субстрата, вовлеченного в этот процесс. Отсюда сделано заключение, что наблюдаемая при выработке условных рефлексов синхронизация электрических потенциалов в разных структурах мозга отражает
Значение функционального состояния лабильности и возбудимости структур, участвующих в образовании временной связи, показано в опытах с созданием искусственных очагов повышенной возбудимости, имитирующих явления
Доминанту рассматривают в качестве основы для формирования временных связей, как переходную форму от суммационного к настоящему условному рефлексу. В доминантном очаге возникают сложные взаимоотношения между нервными и глиальными клетками. Формирование доминанты имеет динамичный характер. Она проходит ряд стадий, среди которых
Образование условных рефлексов, как уже упоминалось, находит отражение в электрической активности не только коры, но и подкорковых структур. При одновременной регистрации потенциалов корковых областей, ядер таламуса и ретикулярной формации среднего мозга в процессе образования условного рефлекса отмечено, что условное возникновение реакции десинхронизации или усвоение ритма сигнального раздражителя может появляться в подкорковых структурах, в частности в ретикулярной формации, раньше, чем в коре (рис. 16), а разрушение неспецифических ядер таламуса устраняет возникшую условную реакцию.
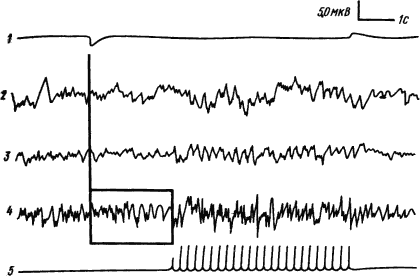 |
Рис. 16. Условно-рефлекторное воспроизведение ритма сигнального раздражения в ретикулярной формации среднего мозга кошки (отмечено квадратом) (по Н. Иошии и др.):
На этом основании было высказано предположение, что временные связи условного рефлекса образуются на подкорковом уровне в неспецифических структурах мозгового ствола. Полагали, что при сочетании условного и безусловного раздражителей по коллатералям их специфических путей происходит конвергенция возбуждений в неспецифические структуры ретикулярной формации среднего мозга. Так объясняли образование их связей, а ослабление восходящих активирующих влияний считали причиной ограничения области десинхронизации в коре. Однако наблюдения за динамикой электрической активности корковых и подкорковых структур в процессе выработки и укрепления условного рефлекса показали, что активность неспецифических образований мозгового ствола наиболее выражена в начальных стадиях выработки, когда они вызывают диффузное возбуждение, а по мере упрочения рефлекса на первый план выступают специфические системы на уровне коры (показано в опытах на кошках и собаках).
Деятельность неспецифических структур мозга, в частности восходящей активирующей системы ретикулярной формации среднего мозга, как показали исследования, действительно играет важную роль в образовании временных связей, обеспечивая высокую степень работоспособности высших отделов мозга. Замыкание временных связей является многоуровневым процессом, в который вовлекаются как кора, так и подкорковые образования, причем большую роль в этих процессах играет гиппокамп.
Широкое участие различных структур мозга в образовании временных связей проявляется и в синхронизации ритмов электрической активности корковых областей и многих подкорковых структур. Однако функция большинства из них, особенно неспецифических областей, связана с созданием условий для образования временных связей. Это достигается активацией высших отделов мозга и распространением этого активного состояния на обширные области коры. По мере укрепления связей эти функции отпадают и участие неспецифических областей становится все более ограниченным. Многие электрические показатели этих процессов, резко выраженные в начале выработки рефлекса, в дальнейшем ослабляются и могут исчезнуть. Так происходит с условной реакцией десинхронизации ЭЭГ человека при упрочении условного двигательного рефлекса. Как видно из рис. 17, четкая депрессия альфа-ритма в отведении, соответствующем корковому участку движения руки, по мере увеличения числа сочетаний становится все меньше и при прочном рефлексе совершенно исчезает.
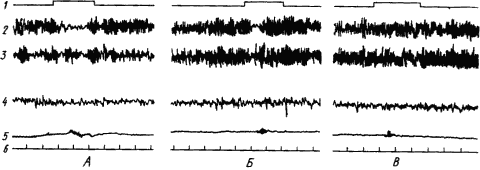 |
Рис. 17. Изменение ЭЭГ человека при выработке условного двигательного рефлекса на звук метронома.
Исследование фоновой ритмики при выработке условных пищевых и оборонительных рефлексов у кроликов в условиях свободного поведения показало учащение колебаний ЭКоГ и укорочение периода условного ответа. При этом наблюдали сходный характер изменений электрической активности при пищевом и оборонительном рефлексах. В специальных опытах с созданием пищевой (голодание) или оборонительной (болевое раздражение) доминанты в коре, ретикулярной формации и лимбических структурах возникали синхронизированные колебания, отражающие высокий уровень неспецифической активации. Эта активация проявлялась не только в учащении импульсов ответов нейрона на условные раздражители, но и в периоды между их предъявлением (рис. 18). Так в деятельности нейронов отражались фазические и тонические компоненты ее перестроек при образовании как пищевых, так и оборонительных условных рефлексов. Однако прямо противоположная реакция в случае пищевого или болевого подкрепления сигнала означает, что проявившаяся в электрической активности неспецифическая активация избирательно действует на структуры мозга, реализующие процессы разного биологического значения. Пищевое подкрепление условного рефлекса придает ему эмоционально-положительный, а оборонительное — эмоционально-отрицательный характер.
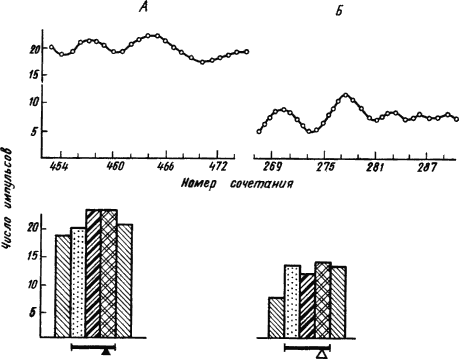 |
Рис. 18. Тонические и фазические импульсные реакции нейронов гиппокампа при условном пищевом
Процессы образования временных связей находят отражение и в
Обнаружение связи медленных электрических процессов с психической деятельностью человека (Н.П. Бехтерева, 1966) было исходным для целого направления исследований их динамики, например при обучении иностранным словам или использовании для анализа нейрофизиологических механизмов мнестической деятельности и процесса формирования матрицы долгосрочной памяти. В последнее время СМКП связывают с метаболическими процессами, влияющими на электродный потенциал, а их сдвиги, наблюдаемые при выработке условного рефлекса, обозначают как колебания биоэлектрохимической активности.
Таким образом,
Вызванные потенциалы. Рассмотренное выше явление усвоения ритмов раздражения представляет собой серии укороченных вызванных потенциалов, в которых полное развитие каждого из них прерывается возникновением следующего. В более полном виде
Так, в первых исследованиях динамики ВП проекционных зон сигнального раздражителя при выработке условного рефлекса одни авторы отмечали его увеличение, другие — уменьшение, третьи — вообще ставили под сомнение наличие закономерной корреляции между ходом образования условного рефлекса и изменениями ВП. Видимо, причина противоречивых результатов состояла в том, что изменения ВП в чрезвычайной степени зависят от стадии образования условного рефлекса (генерализованного или специализированного, непрочного или упроченного), от локализации отведения (проекция условного или безусловного раздражения, первичные или вторичные сенсорные зоны), от вида, возраста и состояния объекта.
Дальнейшие исследования наглядно показали наличие такой зависимости. Изменения ВП происходят по стадиям формирования условного рефлекса. В большинстве случаев начало образования временной связи может сопровождаться некоторым снижением величины ВП. На стадии генерализации условного рефлекса иррадиация возбуждения в коре мозга проявляется в виде усиления и широкого распространения ВП почти на все области коры и укорочения латентных периодов их ответов на раздражители.
На стадий упрочения условного рефлекса происходит ограничение области возникновения ВП, отражающее концентрацию процессов возбуждения в его исходном очаге. При этом величина и форма ВП приближаются к исходным. Обобщение результатов исследования динамики ВП в сопоставлении с другими показателями выработки условных рефлексов позволило заключить, что процесс отражения окружающего мира меняет свой характер на разных этапах условно-рефлекторной деятельности. Например, начальное увеличение с последующим снижением негативной волны ВП соответствует восстановлению угашенной реакции внимания. Возрастание амплитуды негативной волны идет параллельно приобретению условным раздражителем сигнального значения.
Сравнение динамики ВП в разных проекционных зонах коры показало, что его уменьшению в зоне проекции сигнального раздражителя может соответствовать возрастание величины ВП в зоне проекции безусловного, служащего подкреплением. Однако в случае инструментального условного рефлекса отношения усложняются. Так, в зоне проекции конечности, выполняющей инструментальное действие, ВП концентрируются на очень ограниченном участке, но оказываются более выраженными в области коркового представительства сигнала. В начальных стадиях образования временных связей ВП указывают на преобладание активности первичных сенсорных зон, потом возрастает роль вторичных.
Особое значение за последнее время приобретает изучение динамики поздних компонентов ВП, следующих за первичным ответом, его позитивных и негативных медленных волн. Так, при выработке зрительных оборонительных условных рефлексов отмечались закономерные перестройки именно поздних волн ВП на вспышки света. Особенно выраженные изменения происходили в медленной негативной волне. Наряду с изменениями формы и амплитуды основных компонентов ВП появлялись новые, ранее не имевшие места волны потенциалов. Если сигналом для условного оборонительного рефлекса сделать низкочастотное раздражение неспецифических ядер таламуса, то в комплексе корковых потенциалов вовлечения по мере выработки рефлекса формируется дополнительная поверхностно-положительная волна, которая при его угашении постепенно исчезает. В то же время поверхностно-отрицательная волна ВП, регистрируемого в ограниченном участке коркового представительства конечности, совершающей оборонительное движение, оказывается более стабильной. Выработка оборонительного условного рефлекса на свет при подкреплении электрокожным раздражением приводит к тому, что ВП на оба эти стимула становятся сходными по своей форме.
Таким образом,
Реакции нервных клеток. Применение микроэлектродов, погружаемых в нервную ткань, позволило исследовать электрическую активность отдельных нейронов при условно-рефлекторной деятельности. Для этого держатель микроэлектрода с регулятором его погружения укрепляли на голове подопытного животного, например обезьяны. Когда был выработан условный оборонительный рефлекс отдергивания руки на вспышки света, то импульсные ответы корковых нейронов на эти вспышки отличались от ответов до выработки условного рефлекса. Характерно, что изменения импульсных ответов у одних нейронов протекали в сторону усиления разряда, а у других — в сторону его ослабления. В этом проявляется сложная функциональная организация коркового процесса условного возбуждения как динамической мозаики возбужденных и заторможенных нейронов.
Соотношение нейронных реакций, отражающих центральные процессы условного возбуждения, оказывается различным в областях коры, осуществляющих разные функции.
Таблица 1. Распределение возбуждающихся и тормозящихся нейронов в разных корковых областях мозга обезьяны при действии условного светового раздражителя (по Г. Джасперу и др.)
 |
Как видно из табл. 1, в зонах проекции условного раздражителя и представительства двигательной реакции возбуждающихся нейронов зафиксировано примерно втрое больше, чем тормозящихся, а в зонах коры, выполняющих ассоциативные и интегративные функции (теменных и лобных), преобладали тормозящиеся нейроны, которых было почти вдвое больше, чем возбуждающихся. Видимо, для осуществления интегративных функций, более сложных, чем начальная обработка информации, в первичных проекционных зонах возрастает роль торможения, формирующая тонкую мозаику нейронной активности. В опытах на кошках также оказалось, что в областях, не являющихся зонами проекции условного и безусловного раздражителей, преобладают тормозящиеся нейроны. Однако в опытах на кроликах более половины нейронов и в зоне представительства условного слухового раздражителя отвечали на сигнал торможением. Доля нейронов, реагирующих торможением на сигнал, увеличивалась и в первичной соматосенсорной зоне мозга крысы при выработке мигательного рефлекса.
Роль тормозящихся нейронов в процессе образования условного рефлекса может заключаться в ограничении числа действующих синаптических связей, выделении специфических рецептивных полей и, тем самым, выделении необходимой информации из исходного шума. Вместе с тем анализ межнейронных связей в слуховой коре кошки показал, что при выработке условного рефлекса на звук усиливались возбудительные связи.
Характер условных реакций нейронов во многом зависит от организации их
Так, в двигательной области коры кролика «обучающихся» нейронов было 78 % среди бимодальных, т.е. отвечающих на два рода раздражений, и только 34 % среди мономодальных, т.е. отвечающих лишь на раздражения одного рода. В коре мозга кошки 90 % полимодальных нейронов являются «обучающимися».
Способность нейронов включаться в условные реакции в значительной мере зависит от уровня возбудимости, о которой можно судить по частоте их фоновой импульсации. Именно нейроны высокой возбудимости, на которые сходятся сигналы от многих полей рецепции, играют важную роль в механизме образования временных связей. Вероятно поэтому нейронов, проявляющих условные реакции, в первичных проекционных полях оказалось меньше, чем во вторичных, где сопоставляется информация, приходящая по многим путям.
Так, в первичном зрительном поле 17 «обучающихся» нейронов было лишь 18 %, в то время как во вторичном зрительном поле 18 их оказалось 68 %. Вместе с тем «обучающиеся» клетки в большем или меньшем количестве можно найти в любой структуре мозга.
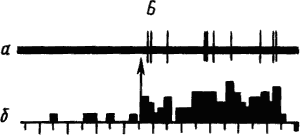
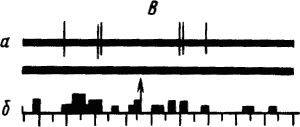
Активность «обучающихся» нейронов изменяется в соответствии со степенью выраженности внешних проявлений условного рефлекса и зависит от факторов, влияющих на эти проявления. Например, насыщение животного пищей ведет к уменьшению проявлений его пищевых условных рефлексов и резкому снижению условных реакций корковых нейронов. При инструментальных условных рефлексах такая зависимость наглядно показана для нейронов лобных отделов коры мозга обезьяны (рис. 19).
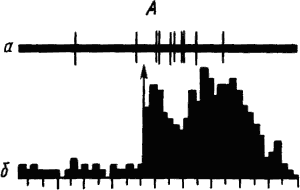 |
 |
 |
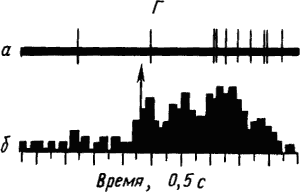 |
Рис. 19. Участие нейрона лобной коры обезьяны в реализации инструментального условного пищевого рефлекса в зависимости от потребности в пище.
Странность, на первый взгляд, условных ответов корковых нейронов на действие сигнального раздражителя состоит в том, что их латентные периоды оказывались более длительными, чем при применении этого раздражителя как индифферентного (до сочетаний с безусловным). Так, некоторые корковые нейроны кошки при выработке оборонительного условного рефлекса на звуковой сигнал могли увеличивать латентный период своего импульсного ответа более чем втрое, от 30 до 100 мс. Для объяснения этого феномена, вероятно, следует исходить из того факта, что включение нейронов в механизм временной связи меняет их положение в нейронных сетях проведения возбуждения и состояние синаптической проводимости. Гистограммный анализ показал, что удлинение латентного периода получено в среднем из сложения удлиненных латентных периодов реакций одних нейронов и укороченных латентных периодов реакций других нейронов. Поэтому можно думать, что при формировании временных связей происходит наряду с усложнением путей передачи сигналов образование нейронных цепочек облегченного их проведения.
На этапе генерализации вырабатываемого условного рефлекса начинают вовлекаться нейроны обширных областей коры и подкорковых структур, гиппокампа, гипоталамуса, ретикулярной формации среднего мозга. По мере укрепления рефлекса реакции нейронов идут на убыль, прежде всего в неспецифических областях и подкорковых структурах, а затем в коре. Специализация условных реакций сопровождается уменьшением числа «обучающихся» клеток, что, видимо, отражает процессы концентрации условного возбуждения. Уменьшаются ответы нейронов как в зоне проекции безусловного, так и условного раздражителей. Однако в специфических подкорковых путях проявления условных реакций сохраняются, в то время как они идут на убыль в корковых зонах проекции условного и безусловного раздражителей.
Например, у кроликов при выработке оборонительного условного рефлекса на звук возникшая вначале условная реакция при дальнейшем укреплении рефлекса практически исчезала у 20–30 % нейронов в слуховой и двигательной зонах коры, хотя в подкорковой области слухового пути (внутреннем коленчатом теле) такую затухающую активность обнаруживали лишь 16 % нейронов. При достаточно длительном многократном повторении сочетаний, обеспечивающем высокую прочность вырабатываемого условного рефлекса у кошек, можно было достичь практически полного возвращения импульсных реакций значительной части нейронов на сигнальный раздражитель к тому уровню, когда этот раздражитель был индифферентным (табл. 2).
Таблица 2. Динамика условных реакций 100 нейронов коры мозга кошки при выработке и упрочении зрительно-двигательного условного рефлекса (по А.Б. Когану)
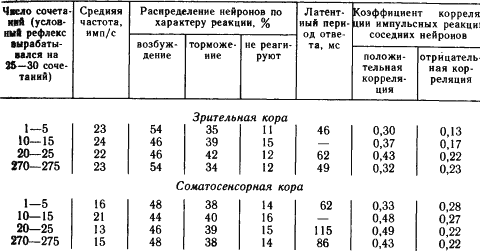 |
Из табл. 2 видно, что такие показатели условных реакций популяций нейронов, как средняя частота импульсов в ответе и его характер при очень прочном укреплении рефлекса, возвращаются в исходное состояние. От них отстают показатели латентных периодов, имеющие еще высокие значения. Наиболее стабильными оказались условные изменения коэффициентов корреляции импульсных разрядов. Обратное развитие условных реакций именно корковых нейронов при упрочении условного рефлекса можно объяснить концентрацией возбуждения, покидающего исследуемые нейроны, или «передачей» автоматизированного навыка на более низкие уровни управления. Однако сохранение при этом показателей корреляции импульсных потоков, установившихся в результате выработки условного рефлекса, указывает на то, что если при формировании временной связи происходят изменения деятельности обширных популяций нейронов, то после завершения процесса его наличие проявляется главным образом в новых сложившихся при этом межнейронных отношениях.
Сопоставление латентных периодов импульсных ответов нейронов и вызванного потенциала показало, что началу первичного ответа соответствует интенсивная активация нейронов в зоне проекции условного раздражителя, к которой присоединяется основная масса реагирующих нейронов в зоне проекции безусловного раздражителя, а более позднему, негативно-позитивному колебанию ВП соответствует тормозная пауза и посттормозный импульсный разряд нейронов.
Исследования нейронной активности мозга человека, проводимые с помощью хронически вживленных с лечебной целью электродов, позволили выявить отражение в этой активности различных форм психической деятельности человека, в том числе процессов обучения (подробнее см. в гл. 17).
Процессы замыкания временных нервных связей при образовании условного рефлекса. Несмотря на многочисленные исследования, до сих пор нет единого мнения о том, что представляют собой процессы замыкания при образовании условного рефлекса, хотя существует много гипотез, каждая из которых подкрепляется солидным фактическим материалом. Одна из причин такой ситуации может состоять в том, что на разных этапах эволюции эти процессы могут иметь различную природу. Так, не вызывает сомнений, что внутриклеточные временные связи простейших, образующиеся при выработке у них примитивных условных реакций, существенно отличаются от межклеточных временных связей в нервной системе. Однако и условные нервные связи далеко не однородны по способу образования, проявлениям и, вероятно, механизмам их осуществления.
Исходя из свойств рефлексов было выделено шесть уровней эволюционного совершенствования их временных связей (Л.Г. Воронин, 1979). Первый уровень —
Естественно предполагать, что при формировании столь различных по своему назначению и устройству механизмов обучения приспособительному поведению могут быть использованы разные способы образования временных связей. Отсюда различие и трудность сопоставления результатов, полученных на разных объектах. Вместе с тем для всех видов временных связей, во всяком случае между нервными клетками, центральным остается вопрос о мембранных механизмах повышения эффективности синапсов и организации нейронных сетей.
За последнее время получено немало фактов о нейронных механизмах условного рефлекса, начинают накапливаться сведения о роли в этих механизмах синаптических и мембранных процессов. Образование временных связей, изучаемое методом условных рефлексов, лежит в основе явлений памяти; этот материал подробно изложен в гл. 3.
| © 2025 Библиотека RealLib.org (support [a t] reallib.org) |