"Дифференциальная психология. Индивидуальные и групповые различия в поведении" - читать интересную книгу автора (Анастази Анна)
Глава 4 НАСЛЕДСТВЕННОСТЬ И ОКРУЖАЮЩАЯ СРЕДА: МЕТОДОЛОГИЯ
Психологи используют самые разные процедуры, пытаясь распутать тот клубок наследственных факторов и факторов окружающей среды, который определяет развитие поведения. Среди них только несколько подходов могут помочь найти вразумительные ответы на возникающие в связи с этим вопросы. У каждого из них есть свои преимущества и свои недостатки. Наше понимание происхождения поведенческих различий может продвинуться, если попытаться объединить результаты, полученные представителями разных направлений.
Главная трудность, с которой сталкивается исследователь в этой области, состоит в том, чтобы выделить действие наследственного фактора и воздействие фактора окружающей среды. Так и при проведении любых экспериментов основной предпосылкой является создание таких условий, в которых можно было бы сравнивать между собой изменяющиеся показатели групп и субгрупп по некоему
Если наследственность одинаковая, как в случае с однояйцевыми близнецами, то различия между индивидами будут являться следствием действия исключительно окружающей среды. Если окружающая среда является одной и той же для разных индивидов, то наблюдаемые различия между ними будут связаны с наследственностью. В свете того, что было сказано в главе 3 о том, что представляет собой окружающая среда, очевидно, насколько сложно сохранять окружающую среду постоянной для двух индивидов, особенно если речь идет о людях. Среди прочих психологи использовали некоторые техники, которые не предполагали попыток разделить наследственные факторы и факторы окружающей среды и, следовательно, могли дать результаты скорее описывающие, чем объясняющие. С другой стороны, есть подходы, которые позволяют в определенном смысле идентифицировать наследственные факторы или факторы окружающей среды, касающиеся индивидуальных различий по некоторым качествам.
Оценивая каждый метод, мы должны держать в уме отмеченную в предыдущей главе специфику изучаемой проблемы. Существенно то, что мы не задаемся вопросом, зависит ли какое-либо качество от наследственности или окружающей среды. Проблема состоит в раскрытии
Почти каждое исследование в дифференциальной психологии может что-то дать для анализа происхождения поведенческих различий. В этом смысле в настоящей главе содержится материал для всех остальных глав данной книги. Существуют определенные техники, которые создавались специально для исследований наследственности и окружающей среды и традиционно ассоциирующиеся в психологии именно с этой проблемой. В последующих разделах мы рассмотрим пять главных подходов. Это: селективное выведение психологических качеств, исследования нормативов развивающегося поведения, исследование структурных факторов в развитии поведения, исследование влияния на поведение предшествующего опыта и статистический анализ родовых сходств и различий.
Экспериментальное селективное выведение животных на основе поведенческих характеристик является современным применением техники селективного выведения, долгое время использовавшейся генетиками. Этот метод генетики является базовым для изучения наследования любых признаков. Создав несколько модификаций этого метода, генетики с успехом проанализировали наследственный базис многих структурных характеристик и даже сконструировали для определенных видов теоретические «генные карты». От селективного выведения до современного применения его к поведенческим феноменам пока далеко. Все, чего на настоящий момент достигли эти исследования, — это появления через последовательное селективное выведение поколений двух наследственных ветвей, существенно отличающихся друг от друга по определенным поведенческим характеристикам. Выводя эти контрастные ветви, исследователи положили начало определению возможных структурных базисов для таких поведенческих различий между ветвями.
Одним из наиболее масштабных исследований с использованием селективного выведения является то, которое провел Трион (85, 86) с белыми крысами, познающими лабиринт. Изначальной группе из 142 крыс дали 19 попыток для прохождения лабиринта, при этом количество «ошибок», то есть попаданий в тупиковые ходы, фиксировалось для каждого животного. На основе результатов для участия в дальнейших экспериментах отбирались сильнейшая и слабейшая группы крыс. Самые «умные» крысы в родительском поколении (Р) спаривались друг с другом, то же самое проделали с самыми «глупыми» крысами. Через эту процедуру провели 18 дочерних поколений (от F до F|S). В каждом следующем поколении самые «умные» крысы в «умных» ветвях отбирались на основе показателей прохождения лабиринта и собирались вместе, самые «глупые» в «глупых» ветвях также отбирались и собирались вместе; внутри каждой из этих групп особи скрещивались. Условия окружающей среды, такие как пища, освещение, температура и жилые помещения, были одними и теми же для всех поколений.
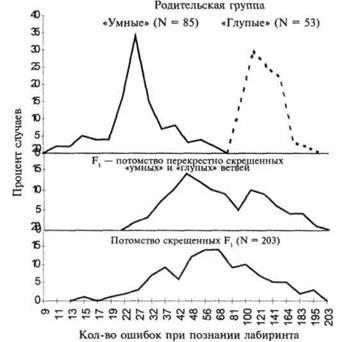
Результаты такого селективного вывода, основанные на показателях прохождения лабиринта каждым последующим поколением, даны на рисунке 26. Необходимо отметить, что распределения «умных» и «глупых» ветвей постепенно расходятся до тех пор, пока на уровне поколения F7 не расходятся совершенно. После седьмого поколения дополнительный эффект от селективного выведения становится незначительным. Когда крысы из «умной» и «глупой» групп были скрещены между собой, распределение результатов стало похоже на распределение результатов родительской группы, — большинство животных показали средние показатели, и лишь некоторые особи выдали крайние значения «ума» и «глупости». Распределения «умных» и «глупых» родительских групп и двух дочерних поколений, полученных в результате скрещивания, даны на рисунке 27.
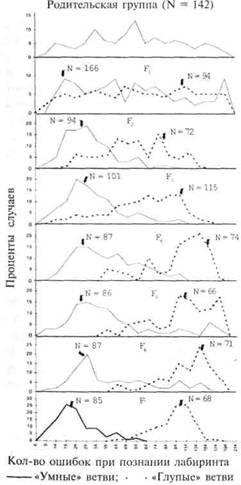 |
Трион сделал вывод, что результаты такого эксперимента по перекрестному скрещиванию подтверждают гипотезу многофакторного наследования. Халл (34) отметил, что если бы ветви оказались беспримесными на момент перекрестного скрещивания, то потомки первого поколения (F,) должны были показывать однообразные значения по прохождению лабиринта, в то время как их собственные потомки (F2) должны были показывать широкий спектр результирующих значений. Тот факт, что поколение F, получившееся в результате перекрестного скрещивания, показало большие индивидуальные различия по данным прохождения лабиринта, означает, что попытка достичь гомозиготности оказалась неудачной и что генетические интерпретации должны носить соответственно более сложный характер.
В своих последующих исследованиях Трион и другие ученые попытались выяснить, чем отличаются «умные» крысы, прошедшие лабиринт с наивысшими показателями, от «глупых» крыс, чьи показатели оказались самыми низкими. Вскоре выяснилось, что такое различие не связано ни с «умственными способностями», ни даже с общей обучаемостью, поскольку крысы, показавшие низшие значения в лабиринте, при решении других задач на обучение показали значения не ниже, а в ряде случаев выше значений тех крыс, которые в лабиринте были самыми «умными». Вслед за Трионом Серл (68) произвел 30 измерений в группах крыс из обеих ветвей. Анализ результатов показал, что крысы, показавшие себя самыми умными в лабиринте, обладали «особенно хорошим аппетитом, склонностью находить кратчайшую дорогу, не особенно боялись воды, были пугливы на открытых пространствах», в то время как крысы, показавшие себя самыми глупыми в лабиринте, были «сравнительно равнодушны к еде, обладали повышенной водобоязнью и пугливостью в отношении механических приборов» (68, с. 323). Таким образом, выяснилось, что представители двух ветвей различались, прежде всего, не своими способностями, но эмоциональными и мотивационными особенностями.
 |
Позже ветви, состоящие из «умных в лабиринте» и «глупых в лабиринте» крыс, успешно скрещивались другими исследователями (37, 43, 71, 83). Им удалось продвинуться в вопросе идентификации структурных различий между ветвями. Состояние желез, размер тела, величина мозга и факторы, связанные со здоровьем, активностью и аппетитом, — все эти признаки зарекомендовали себя в качестве возможного основания различения ветвей крыс, познающих лабиринт. Сравнивание разных ветвей по физическим параметрам их представителей давало иногда противоречивые результаты, — кто-то из исследователей находил существенное отличие по определенному физическому параметру, а кто-то нет. Таких несоответствий можно было бы ожидать, если принять во внимание то, что на активность крыс, подобную познанию лабиринта, оказывает влияние
Селективное выведение проводилось также в отношении таких свойств крыс, как спонтанная активность (3, 63) и эмоциональность (33). Результаты одного из исследований (35) навели на мысль о существовании определенных отношений между свойствами, селективно выведенными, и другими поведенческими характеристиками. Например, сравнительно неэмоциональные, «бесстрашные» крысы показали более высокий уровень активности и большую агрессивность, чем сравнительно эмоциональные.
Чрезвычайно важный шаг в развитии генетики поведения был сделан в исследовании Хирша и Триона, которое они провели на низших организмах (45). Эти исследователи разработали техники, позволяющие проводить надежные измерения индивидуальных различий в поведении таких организмов, как фруктовая муха
Несколько другой подход представляет собой исследование поведенческих различий между существующими ветвями животных, которые не были селективно выведены для получения психологических свойств. Одно такое исследование трех, внутри себя скрещивающихся мышиных ветвей показало существенные отличия между ветвями по признаку агрессивности (64). Несколько исследовательских проектов этого типа были реализованы Стокардом с собаками (77), а после него — Скоттом и Фулером в Лаборатории имени Джексона (25, 65, 66, 67). Различные перекрестные скрещивания собак приводили к существенным различиям в количестве доступных для решения собаками задач, так же как и в эмоциональных реакциях, уровне активности и в социальном поведении, проявляемом по отношению к другим собакам и к людям из числа обслуживающего персонала. Как и при исследовании крыс, снова проявилось то, что различия в способностях к обучению «связаны с различиями в эмоциональных, мотивационных и периферических процессах и что генетически обусловленные различия в центральных процессах могут быть несущественными или не существовать вовсе» (66, с. 225). В ходе исследований собак были также обнаружены определенные, полученные в результате скрещиваний различия в психологических свойствах, которые, в свою очередь, были связаны с наблюдаемыми поведенческими различиями.
Каких бы животных видов ни касались исследования, в большинстве случаев наиболее вероятным источником поведенческих различий, наблюдаемых в наследственных ветвях, было функционирование эндокринных желез. Что же касается наследственных различий в свойствах мозга или других частях нервной системы, то этот вопрос остается открытым по сей день (ср. 34). При интерпретации результатов экспериментов по селективному выводу необходимо учитывать следующую методологическую трудность, замеченную Холлом: «Передача какого-либо свойства от поколения к поколению может происходить генетически, но в реальности это часто происходит благодаря действию другого фактора. Эта передача может являться результатом пренатального или постнатального влияния матери на потомка. Пренатальное влияние может быть связано с трансплантацией оплодотворенной яйцеклетки матери из другой наследственной ветви. Постнатальное влияние можно изучать, поместив новорожденных к приемной матери» (34, с. 309). На настоящий момент очень мало исследователей пытались осуществить это на практике. В то же время существуют серьезные свидетельства (они будут рассматриваться в следующем разделе), которые показывают, что все формы поведения в условиях селективного выведения за счет оказываемого в раннем возрасте влияния окружающей среды, имеют различную изменяемость.
Другой источник данных, касающихся этиологических факторов в поведении, можно найти в наблюдениях за нормальным процессом развития поведения. Какую информацию о взаимодействии фактора наследственности с фактором окружающей среды может дать такой подход? Большинство исследователей, использующих его, ищут свидетельств
Анализируя данные нормального развития с точки зрения гипотезы созревания, чаще всего ссылаются на два обстоятельства:
Перед нами огромный массив данных наблюдения за поведенческим развитием самых разных форм жизни — от саламандры до человека. Относительная простота процессов у низших форм облегчает понимание основных принципов развития и их применение к высшим формам. Подобным образом, некоторые из наиболее существенных наблюдений были сделаны на пренатальном уровне потому, что поведение на этом уровне гораздо проще, а также потому, что разнообразие окружающей среды не так велико, как в течение постнатальной жизни. Естественно, процесс поведенческого развития продолжается в течение всей жизни, и включает в себя, характерное для старости, ухудшение функционирования. Поскольку возрастные изменения у детей старшего возраста и у взрослых мы будем рассматривать в одной из следующих глав (гл. 8), сейчас мы рассмотрим то, что происходит во время пренатального периода[14]. Полный обзор исследований пренатального поведенческого развития, как у человека, так и у человекообразных видов, можно найти у Кармихаэля (9). В кратком варианте его изложил Манн (60, гл. 6). В этих работах содержится также введение в специализированные техники, которые были разработаны для наблюдений пренатального поведения у человека и животных и критическая оценка таких процедур. Среди классических исследований в этой области можно упомянуть исследование саламандр Кохиллом (10), кошек Корониусом (11), цыплят Куо (52, 53, 54, 55) и гвинейских свиней Кармихаэлем (7). Выдающийся вклад в исследование этой темы недавно был сделан Хукером (47), который снял фильм о поведении человеческого плода.
В исследованиях постнатального поведенческого развития животных активное участие принимал Крюкшанк (12). Пратт (61) обобщил наблюдения за человеческими новорожденными. Изучение поведенческого развития у новорожденных и детей относится к предмету детской психологии. Результаты исследований в этой области опубликованы в многочисленных работах. Гессель и его сотрудники в клинике Детского развития Йельского университета (ср. 28, 29, 30) наиболее эффективно проанализировали данные по детскому развитию с точки зрения гипотезы созревания. За годы своей работы этой группой ученых было собрано большое количество данных по нормальным циклам развития в младенчестве и детстве. На основании этих данных Гессель и его сотрудники создали нормативный каталог, показывающий для какого возраста характерны какие функциональные изменения. Их наблюдения основывались на тщательно разработанных процедурах, предполагающих использование стандартизированных игрушек, кроваток, стульчиков и другого оборудования, экранов, прозрачных с одной стороны, киносъемки и других приспособлений.
По всем исследованным видам были получены четкие свидетельства о том, что поведение возникает задолго до рождения. У человеческого плода, например, реакции на тактильные прикосновения были зафиксированы уже через восемь недель после зачатия. Считается, что все рецепторы способны функционировать до рождения, хотя зрение, вкус, обоняние и температурные ощущения вряд ли стимулируются вследствие однообразия условий пренатальной окружающей среды. У человеческого восьминедельного плода были зафиксированы несколько типов моторных реакций. На четвертом месяце беременности проявляются почти все рефлексы, которыми обладает новорожденный. Более того, многие реакции человеческого плода являются предвестниками поведения в последующей жизни. Среди них такие рефлексы, как рефлекс плача, сосательный и глазной рефлексы; рефлекс равновесия и ориентировки; рефлекс ритмичных сокращений грудной клетки, напоминающих дыхательные, и рефлекс «перебирания ножками», заключающийся в сгибании и разведении конечностей в разные стороны. Считается, что этот рефлекс перебирания ножками лежит в основе таких постнатальных действий, как ползание и ходьба. Существуют также свидетельства того, что неродившийся еще ребенок уже способен к элементарному обучению, хотя исследователи считают, что даже родившийся ребенок в первый месяц своей жизни не может правильно воспринимать окружающий мир (ср. 8, 19, 74, 88).
Пренатальное поведение было также зафиксировано у многих животных. Существование обусловленных реакций было установлено у эмбрионов птенцов (32, 48). Проводя многочисленные серии наблюдений за поведением эмбриона птенца, Куо (54) обратил внимание на механическую и иную стимуляцию, на которую реагирует эмбрион, а также на многочисленные типы мускульных реакций, которые нарабатываются до рождения. Корониус (11) на основании проведенного исследования кошек сделал следующий вывод: «До рождения существует развитие поведения, — быстрое, прогрессирующее и продолжительное… «Примитивные» реакции, связанные с дыханием, ориентировкой, локомоцией и кормлением, являются результатом длительного и прогрессирующего пути пренатального развития» (11, ее. 377–378). Эти открытия являются предостережением против того, чтобы принимать «внезапное появление» поведенческих функций в качестве критерия созревания. Внезапное появление определенного действия после рождения может в некоторых случаях представлять собой при условии наличия подходящего стимулятора в окружающей среде повторное появление поведения, которое уже прошло путь постепенного развития и наработок до рождения.
Изучение развития раннего поведения позволило получить многочисленные свидетельства последовательного проявления моделей поведения. Единообразие хода развития и порядка появления поведенческих изменений, как видно, является правилом для функций, возникающих во время пренатального и раннего постнатального периодов. Изучение животных позволило разработать подробные «временные таблицы» поведенческих изменений, по которым более или менее точно можно определять то, как будут развиваться представители каждого вида. Такая же информация была собрана в отношении развития человеческого плода.
Сходным образом на Гесселя и других детских психологов произвела впечатление повторяемость последовательности развития поведения в младенчестве и раннем возрасте, которой они дали объяснение с позиций гипотезы созревания (ср. 1, 21, 29, 30, 36, 69, 70). Для хватательного поведения, например, последовательность этапов его развития у разных детей для соответствующих возрастов приблизительно одна и та же. Так, реакции детей на маленький сладкий шарик, лежащий перед ними, показывают характерную хронологическую последовательность, связанную с движениями глаз, рук и пальцев. Использование всей руки в неумелых попытках «захвата ладонью», например, наблюдается в более раннем возрасте, чем использование отставленного от ладони большого пальца; после этого следует использование большого и указательного пальцев в процессе осуществления более эффективного «пинцетного» захвата шарика. Такое последовательное проявление моделей поведения зафиксировано в развитии ходьбы, залезания на лестницу и большинства видов сенсомоторного развития в первые годы жизни.
В подтверждение гипотезы созревания Гессель (30) ссылается на преждевременные роды и роды, происшедшие с задержкой, то есть на такие роды, которые произошли раньше или позже нормального девятимесячного срока беременности. Как соотносится поведение одномесячного младенца, родившегося преждевременно, в восемь месяцев, с поведением нормального ребенка, родившегося в срок? Возраст от зачатия у них один и тот же. Но преждевременно родившийся ребенок имеет преимущество в один месяц, — с точки зрения опыта взаимодействия с окружающей постнатальной средой, с ее изменчивыми стимуляциями и широкими возможностями для наработки различных функций. Гессель утверждает, что, несмотря на внешние и связанные со временем преимущества преждевременно родившегося ребенка, матричная модель поведенческого развития у обоих одна и та же.
Короче говоря, получается, что созревание играет главную роль в поведенческом развитии. При этом важно принять в расчет то, что связано с интерпретацией данных в этой области и с квалификацией предложенных обобщений. Во-первых, необходимо обратить внимание на существование
И наконец, важно осознавать
Поскольку любое влияние, которое наследственность оказывает на поведение, как было сказано в главе 3, должно быть проявлено через «структурные» условия, непосредственное изучение роли структурных факторов в развитии поведения представляет собой особый подход к анализу взаимодействий наследственности и окружающей среды. Этот подход часто используют совместно с одной из тех методик исследования, которые мы уже рассматривали. Мы видели, например, как ученые пытались сопоставить
Подобным образом множество исследований нормативов развивающегося поведения включают в себя наблюдения за
Информация, касающаяся изменений в нервной системе, являющихся основанием для развития человеческого поведения в раннем возрасте, была получена через гистологические исследования человеческого плода после смерти. Например, изучение не способного передвигаться плода показало, что в таких случаях в нервной системе еще отсутствовали сенсомоторные связи. Проведя различные процедуры, ученые получили необходимые данные, показывающие, что контроль за поведением со стороны коры головного мозга не возникает раньше, чем через некоторое время после рождения. Электроэнцефалограммы[15] человеческого плода в утробе матери, например, показывают его малую мозговую активность или ее полное отсутствие до момента рождения (57).
Даже когда в нервной системе имеются сильные анатомические взаимосвязи, в появлении соответствующих функций часто наблюдается отставание. Были организованы исследования, направленные на открытие в нервной системе дополнительных факторов, реализация которых предшествует появлению соответствующих функций. Одно время считалось, что существенной обусловливающей предпосылкой является миелинизация, или наличие жировой миелиновой оболочки, окружающей нервные волокна. Вначале исследования показали наличие соответствия между первым появлением специфических функций поведения и миелинизацией собственно нервных путей. Однако эти открытия не получили подтверждения в последующих исследованиях, и сейчас признано, что миелинизация не является предпосылкой функционирования нервных волокон (ср. 75, с. 267).
Находки, связанные с миелинизацией, проливают свет на важный пункт, касающийся интерпретации соответствия структурного и поведенческого развития. Миелинизация может улучшить функционирование, или, наоборот, сама стимулироваться и ускоряться развитием функций, или же два процесса могут протекать одновременно, не влияя друг на друга. То, что корреляция не означает наличия причинной связи, не является трюизмом. Об этом необходимо помнить при оценке роли структурных факторов в поведении. В настоящее время продолжаются поиски возможных гистологических, химических или физических изменений, которые бы предваряли функционирование нервов, но определенного результата пока нет.
Еще одной методикой из числа основных является методика исследования отношения структуры к поведению, состоящая в
Эти исследования психофизиологов, безусловно, явились большим вкладом в наши познания о том, как действуют различные поведенческие структуры. Тем не менее этот подход не слишком сильно продвинул нас в понимании источников индивидуальных различий в поведении в пределах их нормального распределения. Знание структурных предпосылок поведения не обязательно означает то, что есть объяснение индивидуальных различий между теми, кто имеет эти предпосылки. Это все равно, как если бы мы старались открыть причины индивидуальных различий в скорости и стиле ходьбы, исследуя одноногого человека.
С точки зрения индивидуальных различий более перспективным вариантом этого подхода является тот, при котором вызываются физические изменения на ранней стадии развития. В некоторых из исследований этой категории использовались условия, не слишком отличавшиеся от тех, которые действуют в процессе естественного развития. Среди факторов, впоследствии оказывающих влияние на поведение на пренатальном уровне, были исследованы: аноксия (кислородное голодание), воздействие различных химических веществ, недостаточное питание, эндокринная секреция и радиация. В ходе недавних исследований, например, было обнаружено снижение обучаемости крыс, чьи матери во время беременности подверглись облучению X-лучами (26, 56, 81). Эти эффекты не надо смешивать с рассматривавшимся в главе 3 производством при помощи радиации генных мутаций. Мутации передаются по наследству, эффекты, о которых идет речь, не передаются.
И наконец, необходимо упомянуть
Роль физических условий в поведении может находиться в пределах от неизлечимой неврологической или метаболической недостаточности, накладывающей жесткие границы на развитие поведения, до простых физических различий между людьми, что отражается в социальной практике, предлагающей лицам с определенными физическими качествами различное лечение. И то и другое может воздействовать на поведение, но по-разному. Другой вопрос касается способа, посредством которого наследственный фактор и фактор окружающей среды взаимодействуют друг с другом, приводя к появлению рассматриваемых физических качеств. Здесь также есть возможность того, что рассматриваемые физические и психологические качества не находятся друг с другом в причинной взаимосвязи, но что на них обоих воздействует третий фактор, такой, например, как социоэкономический уровень. Так, дети в слаборазвитых регионах могут иметь физические недостатки, являющиеся следствием плохого питания и отсутствия медицинской помощи, и проблемы с интеллектом, связанные с недостатками школьного образования (или отсутствием оного).
Одним из наиболее эффективных способов изучения роли созревания и научения в поведенческом развитии заключается в систематическом изменении первоначального опыта индивида и наблюдении за изменениями его последующего поведения. Большая часть исследований, проведенных с помощью данного метода, была связана с изучением молодых животных. Несколько сопоставимых исследований были проведены на младенцах и детях младшего возраста. Соответствующие данные также были собраны путем сравнения индивидов или групп, подобранных с точки зрения разных критериев, даже если при этом экспериментатор не учитывал различий окружающей среды. Эти исследования представляют собой вариант применения того же основного метода, хотя и предполагающего меньшую контролируемость.
Эксперименты с животными. Множество поведенческих функций, рассматриваемых не как приобретенные в результате научения, а как «инстинктивные», были поставлены под экспериментальный контроль, в результате чего животным не давали нарабатывать определенную функцию до тех пор, пока она не проявлялась сама в соответствующем для данного вида возрасте. Тем самым делалась попытка определить степень, в которой физическое созревание необходимых структур само по себе вело к проявлению данной функции. Другие эксперименты преследовали противоположную цель — обеспечение дополнительной стимуляции данной функции или интенсивной ее наработки для того, чтобы определить, насколько можно ускорить или изменить процесс нормального поведенческого развития.
В 1954 году Бич и Джейнс (2) сделали обзор множества исследований, предшествовавших тем, о которых идет речь. С тех пор интерес ученых к данной теме нисколько не уменьшился, о чем свидетельствуют публикации результатов новых исследований. Среди исследованных вышеуказанным способом функций: полет и пение птиц, плавание лягушек, реакции кормления, запасания пищи, репродуктивное поведение, родительское поведение, самоочищение, кормление молодняка, формирование групп, формирование предпочтений к представителям своего или других видов, реакции восприятия разными органами чувств, борьба и агрессивное поведение, дикость, робость и другие эмоциональные реакции. Все эти функции показывают некие изменения, происшедшие в результате введения экспериментальных условий, хотя степень этих изменений зависит от вида, от функции и от экспериментальных условий.
В некоторых ситуациях типичная для данного вида реакция не проявляется. В других случаях реакция может видоизменяться. Или же реакция на стимул может быть не той, которую он должен был вызывать. Примером последнего является реакция, которую Лоренц и его сотрудники назвали «импринтингом» (ср. 2; 84, с. 150). Это такая реакция, которая, будучи пробуждена у молодого животного первым стимулом, в дальнейшем вызывается только им одним. Так, когда новорожденных гусят изолировали от взрослых гусей и позволили им в течение некоторого времени следовать за экспериментатором, они продолжали следовать за ним и после этого, не проявляя никакого желания следовать за взрослыми гусями. Точно так же птенцы нескольких видов птиц способны воспитываться у птиц другого вида, которые становятся для них «приемными родителями».
Несколько показательных экспериментов помогут нам дать представление о многообразии использованных процедур. Круз (13) изучал реакции кормления у птенцов, которым не давали клевать в течение различных временных промежутков. Пять равных групп, по 25 или 26 птенцов каждая, держали в темноте, а птенцов, являющихся их членами, кормили из руки соответственно в течение одного, двух, трех, четырех и пяти дней после рождения. Затем им позволяли клевать самостоятельно и результаты самостоятельного кормления представлены на рисунке 28. Заметим, что к началу самостоятельного кормления члены всех групп, независимо от возраста, были примерно в равной степени голодны. Как бы то ни было, члены старших групп развивались быстрее и за несколько дней перегнали тех, кто начал самостоятельно кормиться раньше. Такие результаты показывают ту роль, которую играет созревание, поскольку птенцы, достигшие более высокого уровня физического развития, способны были обучаться лучше, чем это делали более молодые птенцы.
Во второй части рисунка 28 график научения группы А, членам которой позволили свободно практиковаться в самостоятельном кормлении после первого дня, сравнивается с аналогичными графиками других четырех групп, которым так же позволили самостоятельно клевать после первого дня, но при этом ограничили их во времени. Членам группы F позволили иметь только 25 сеансов самостоятельного кормления в день, а членам группы G — всего 12 перед тем, как их возвращали в темное помещение. В результате члены обеих групп показали более низкий уровень развития, чем члены группы А. Действительно, члены группы G после первого дня почти остановились в своем развитии. Членам последней группы (Н) позволили иметь 12 ежедневных сеансов самостоятельного кормления в течение первых 10 дней, а после этого по 25 ежедневных сеансов. В течение первых десяти дней результаты группы Н были неотличимы от результатов группы G, хотя группа Н начала быстро опережать группу G, когда ей увеличили ежедневную практику. Таким образом, данный эксперимент показывает, что в поведении птенцов при проведении нормального кормления решающую роль играет совместное влияние созревания и научения.
 |
А. Реакции глотания у членов пяти групп птенцов, которым не позволяли клевать соответственно в течение 1, 2, 3, 4 и 5 дней
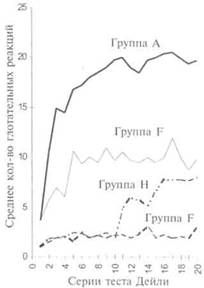 |
В. Реакции глотания у членов четырех групп птенцов, которым позволяли клевать ежедневно в течение разных периодов времени
В аналогичном эксперименте Кармихаэля (4, 5, 6) группу плавающих головастиков, которых после рождения держали в воде с химическими добавками, сравнивали с контрольной группой головастиков, оставшихся в чистой воде. Эффективность химических добавок заключалась в том, что они не позволяли головастикам совершать плавательные движения до достижения ими возраста, в котором они способны нормально плавать. Когда после этого группу головастиков перемещали в чистую воду, они, как только проходило действие химических добавок, начинали нормально плавать. Таким образом, было доказано, что созревание играет большую роль в становлении плавательного поведения таких видов животных. В более позднем эксперименте другому исследователю (24) удалось продемонстрировать существенное преимущество контрольной группы плавающих животных в скорости и расстоянии. Навык, таким образом, играет значительную роль в таком функционировании, хотя базовая моторная модель появляется даже в условиях отсутствия предварительной активности.
В экспериментах, рассматриваемого нами типа, часто трудно быть уверенным до конца, что влияние предварительного научения было действительно устранено. В реальности существуют сложные поведенческие модели, которые обычно называют «инстинктами». Например, изолированная самка крысы, тем не менее, может «строить гнездо» и заботиться о своем потомстве в соответствии с нормами своего вида. Более тщательное исследование, однако, позволяет сделать предположение, что для проявления материнского поведения необходим первоначальный опыт обращения с материалами и опыт ухаживания за своим телом (ср. 62). Самки крыс, содержавшиеся в клетках, в которых не было ничего, из чего можно было бы сконструировать гнездо, впоследствии не могли строить гнезда, когда материал для строительства становился для них доступен. Они не могли также собирать в одно место свое потомство так, как это обычно происходит у крыс. В другой группе самкам крыс надели специальные резиновые воротники, которые не позволяли им прикасаться носом или ртом к нижней части своего тела. Воротники они носили с момента рождения до возраста половой зрелости. Произведя на свет потомство, они не чистили его.
Вместо того чтобы ограничивать активность животных, некоторые исследователи усиленно стимулировали ее, создавая такие условия для развития функций, которые обычно у животных отсутствуют. К этой категории относятся эксперименты с шимпанзе, помещенными в условия жизни людей. В этих случаях к шимпанзе не относились как к домашнему животному, но обращались с ним как с ребенком — во всем, что касалось одежды, условий сна и питания, игр и других аспектов занятий с детьми. Кроме того, вместе с шимпанзе жили собственные дети экспериментаторов, и с шимпанзе обращались по возможности так же, как с детьми. Первыми психологами, осуществлявшими такой проект систематически и целенаправленно, были Келлогсы (50), которые привезли самку шимпанзе по имени Гуа в возрасте семи с половиной месяцев в свой дом и оставили ее там жить на девятимесячный срок.
После этого Хейсы (38, 39, 40, 41) реализовали похожий проект, но по продолжительности более длительный. Вики, другую самку шимпанзе, взяли в дом, когда ей было всего несколько дней от роду; она жила в доме экспериментаторов до самой своей смерти, которая наступила от вирусной инфекции через шесть с половиной лет. Вики вела себя сообразно своему возрасту, так, как ведут себя хорошие дети, оставаясь при этом превосходным лабораторным шимпанзе, чем помогала изучать проблему имитации. Она могла по команде: «Делай это», — повторять действия экспериментаторов, а также спонтанно имитировать выполнение нескольких домашних дел, таких как очинка карандашей или открывание банки консервов. Кроме этого, Вики принимала участие в спонтанных играх с механическими игрушками, причем управлялась с ними не хуже игравших вместе с ней детей; столь же умело Вики обращалась с различными инструментами. По результатам невербальных тестов на умственное развитие, связанных с умением оперировать формами досок, блоками, решать рисованные загадки и т.п., ее развитие в определенном возрасте соответствовало нормам развития человека.
Единственное серьезное затруднение Вики было связано с тем, чтобы овладеть языком. С неимоверными усилиями экспериментаторам в конце концов удалось научить ее говорить три слова (мама, папа, чашка) и использовать их по назначению. При осуществлении функций социального общения Вики предпочитала указывать на интересующий ее предмет, подводить к нему или производить имитирующие движения. Только тогда, когда ничего не помогало, она прибегала к помощи своего голоса. Экспериментаторы сделали вывод, что главная причина трудностей заключалась в неприспособленности речевого аппарата шимпанзе для голосовой передачи слов. У шимпанзе почти полностью отсутствовало столь характерное для человеческих младенцев свойство лепетания, хотя у шимпанзе есть физические возможности воспроизводить широкий спектр речевых звуков. Кроме того, хотя Вики и использовала звуки для выражения эмоций, произвольная вокализация для нее всегда представляла сложность.
Группа ученых, экспериментирующих с животными и занимающихся исследованиями в области дифференциальной психологии, заинтересовалась влиянием, которое оказывает детский опыт на последующие способности научения. В упоминавшемся выше обзоре Бич и Джейнс пишут: «Многие психологи убеждены в том, что способности к научению у животных генетически определены и сравнительно неизменяемы, но недавние открытия показывают необоснованность этого утверждения» (2, с. 255). Многие исследования, проводимые на обезьянах и крысах, свидетельствуют о том, что животные могут «учиться тому, как учиться», иными словами, они могут «обучаться».
Множество современных исследований по этой теме были связаны с теорией, предложенной Хеббом (42), заключающейся в том, что животные, получившие на раннем этапе жизни большой сенсорный опыт, обладают более высокими способностями к научению, чем животные, которые были лишены такого опыта. Необходимые данные, подтверждающие этот тезис, были собраны в ходе исследований крыс и собак (23, 58, 82, ср. также 2). Во всех этих случаях животные, содержавшиеся в менее ограничивающих условиях, показали значительно более высокие результаты в лабиринте и при решении задач.
Эксперименты с младенцами. Эксперименты, связанные с негативным воздействием на нормальное поведение младенцев, должны быть для них безвредны. В исследовании Денниса (16) последствий
На протяжении 14 месяцев поведенческое развитие этих двух детей сравнивалось с нормами, установленными для детей, растущих в обычных, неограничиваемых условиях. Функции, которые обычно появляются в течение первых семи месяцев жизни, у девочек появились без существенной задержки. Среди них были такие простые формы активности, как фиксация объектов; вздрагивание, поворот головы или плач как реакция на звук; захват предметов; наблюдение за своими руками и игра с ними; поднесение руки или предмета ко рту. Очевидно, что такие функции появляются независимо от научения тогда, когда завершается необходимое структурное развитие. Что касается реакций, которые обычно возникают по достижении семимесячного возраста, то они у девочек стали появляться с существенной задержкой. Однако, как только были сняты соответствующие ограничения, эти реакции быстро появились[16].
Техника, которая нашла самое широкое применение в исследованиях детей, называется
Что касается функции лазания, то здесь отличие было зафиксировано. В то время как Т оказалась достаточно опытным верхолазом, ее сестра не могла добраться до верхней ступеньки даже с посторонней помощью. Однако две недели спустя, в течение которых девочка-близнец К по-прежнему не тренировалась, она оказалась в состоянии залезть наверх безо всякой помощи. В этом возрасте (53 недели) девочка-близнец К начала двухнедельный тренировочный период, в конце которого она по умению лазать приблизилась к Т. Таким образом, за счет более выраженного действия эффекта созревания двухнедельный период тренировок в возрасте 53 недель оказался почти столь же эффективным, как шестинедельный период тренировок в возрасте 46 недель.
Та же самая пара однояйцевых близнецов так же последовательно прошла через тренировку других функций, включая словарную подготовку (44, 80). Начиная с 84-недельного возраста у близнеца Т начались интенсивные, длившиеся пять недель занятия, заключавшиеся в изучении названий предметов, выполнении простых заданий и других словарных наработках. Близнец К была лишена всякой возможности слышать в течение этого периода устную речь. По окончании этого пятинедельного периода, когда близнецам было 89 недель, К прошла курс подобных занятий, но он продолжался всего четыре недели. По их окончании словарный запас близнеца К составлял 30 слов. Словарный запас близнеца Т после четырех недель занятий составлял 23 слова, а на конец всего пятинедельного периода составил 35 слов. Исследователи, интерпретируя эти результаты, выделяют роль созревания, указывая на тот факт, что близнец, с которым начали заниматься языком в возрасте 89 недель, почти каждый день продвигался вперед более быстрыми темпами и на каждом этапе занятий демонстрировал более зрелые знания, чем близнец, с которым начали заниматься языком в возрасте 84 недель.
Делая выводы, необходимо отметить, во-первых, что различия в результатах обучения у двух близнецов были небольшими. Например, словарный запас К после 27 дней занятий был равен словарному запасу Т после 31 дня занятий (а именно, 29 слов). Во-вторых, по своему словарному запасу, произношению и конструированию предложений близнец, прошедший пятинедельный курс обучения в более раннем возрасте, немного превосходил другого, прошедшего четырехнедельное обучение в более старшем возрасте, и это превосходство, спустя три месяца, почти сошло на нет. И наконец, очевидно, что развитость структуры в младенчестве и раннем детстве, облегчает формирование первичных языковых навыков. Ребенок не может воспроизводить нужные последовательности звуков, пока его слуховой и голосовой аппарат не позволяет ему осуществлять необходимую их дифференциацию и контроль. Таким образом, факторы, связанные с созреванием, могут влиять на чисто голосовой аспект развития языка, в то время как его символический и смысловой аспекты зависят от обучения.
Интерпретируя результаты всех экспериментов, проведенных с помощью метода близнецового контроля, мы приходим к тому, что на самом деле в них сравниваются не обучение и созревание. Сравниваются, с одной стороны, особый период контролируемой подготовки и, с другой стороны, совокупное действие фактора созревания и всех неконтролируемых наработок определенных функций, осуществлявшихся вне стен лаборатории. Эти неконтролируемые наработки продолжаются до тех пор, пока не предпринимаются блокирующие их меры. Чем больше у близнецов возможностей для таких неконтролируемых наработок, тем меньше различий в лабораторно тестируемых функциях. Поэтому, несмотря на свою популярность, нам представляется, что метод близнецового контроля, строго говоря, не позволяет распознавать факторы, влияющие на развитие личности.
Исследования случаев «диких детей». Широко известные случаи, когда были найдены дети, жившие по большей части в изоляции от людей или в окружении животных, можно рассматривать как «естественные эксперименты» на ограниченное научение. Описания таких «диких детей» встречались в истории давно. В 1758 году Линней включил их в свою классификацию типов человека под названием «человек свирепый». Обзор записанных случаев такого рода был сделан Зингом (89, 90). Ему удалось собрать свыше сорока случаев, правда, информации по многим из них совсем немного и не во всех из них имела место полная изоляция ребенка от людей. Некоторые из этих детей, заблудившись, смогли выжить благодаря своим собственным усилиям, другие воспитывались у таких животных, как волки, медведи, козлы, овцы, буйволы и леопарды.
Дети, жившие в условиях, едва достаточных для выживания, и не имевшие никаких контактов с людьми, были также включены в эту категорию.
Одним из наиболее изученных случаев является случай с Виктором, диким мальчиком Авейрона (49). В сентябре 1799 года во Французском лесу нашли мальчика 11 или 12 лет. Мальчик был полностью обнажен, нечесан, напуган, не мог говорить — одним словом, впечатление было такое, что он вел дикое существование, подобное животному. Он был пойман мужчинами, когда, убегая от них, залез на дерево; его вернули в цивилизацию, где он в конце концов попал в распоряжение и под присмотр французского врача Итарда. Последний опубликовал об этом случае детальный отчет, в котором содержалось как описание первоначальных реакций мальчика, так и история пятилетних занятий с ним.
В более поздние годы были найдены «волчьи дети» Миднапура (27, 50, 72, 76, 79, 89), случай которых был также вполне достоверен. В 1921 году две девочки в возрасте, соответствующем 2–4 и 8–9 годам, были найдены, живущими в пещере с волками в малонаселенном индийском регионе. Их взяли в местный сиротский приют, в котором были предприняты попытки воспитать их. Подробный дневник их поведения, который вел ректор приюта, был опубликован (72), в него вошли анализ и комментарии, сделанные несколькими психологами, социологами, генетиками и антропологами. Было очень трудно поддерживать хорошее самочувствие девочек, в частности, потому, что нормальная человеческая еда вызывала у них слабость и сильные аллергические реакции на коже. Младшая девочка, Амала, не прожила и года; старшая, Камала, жила в течение восьми лет, в течение которых удалось достичь определенного прогресса в обучении ее языку и адаптации ее к человеческому образу жизни.
Следует упомянуть также знаменитый и таинственный случай с Каспаром Хаузером (ср. 72, ее. 277–365), о котором писали очень много. Некоторые соображения позволяют предполагать, что он был наследником царского дома и был устранен политическими противниками. С раннего детства он был заключен в темную камеру, в которой он не мог даже выпрямиться. На нем не было никакой одежды, кроме рубашки и брюк. Когда он просыпался, то находил хлеб и воду, но он не видел того, кто приносил их, и у него не было ни малейшего представления о том, что кроме него в мире еще кто-то есть. Его освободили в 1928 году в возрасте примерно 17 лет. Он бесцельно бродил по улицам Нюрнберга. Он не разговаривал, но время от времени бессмысленно произносил какие-то фразы. Считается, что у него было отличное обоняние и удивительная способность видеть в темноте. Его ходьба напоминала первые шаги ребенка. После различных превратностей судьбы его воспитание взял на себя умелый и терпеливый учитель. В ходе обучения Каспар Хаузер делал успехи и вскоре вновь научился говорить. Он рассказал все, что помнил о своей жизни в камере, а также изложил свои впечатления, связанные с периодом обучения. В отличие от других случаев, когда дети обретали контакт с цивилизацией слишком поздно, Каспар Хаузер смог получить образование, достаточное для того, чтобы достичь уровня обычного человека и даже превзойти его.
Случаи с дикими детьми, подобные этим, являются предметом особого интереса психологов, потому что они могут помочь пролить свет на вопрос, насколько нормальное человеческое поведение может развиваться при отсутствии нормальной стимуляции. Делая выводы из записанных случаев, Зинг (89) сделал заключение, что такие дикие дети, все без исключения, не умели говорить и передвигались на четвереньках. При таких обстоятельствах не могла развиваться ни речь, сколько-нибудь похожая на человеческую, ни специфическое человеческое прямохождение. У всех у них развилась форма передвижения на руках и ногах, или на руках и коленях, а их физическая структура при этом, как правило, модифицировалась для осуществления быстрого и эффективного передвижения на четырех конечностях (на руках и ногах появились мозолистые подушечки и т.д.).
Характерные модификации, как сообщается, касались и их органов чувств: обоняния, слуха, зрения, — которые обрели звериную остроту. То, как они употребляли пищу, было нетипичным для людей. Сырое мясо являлось обычной пищей детей, воспитывавшихся плотоядными животными; дикие дети, жившие самостоятельно, существовали, как сообщают, за счет употребления коры деревьев, кореньев, травы, растений и листьев. Одна из диких девочек во Франции приспособилась ловить в воде рыбу и лягушек, которые составляли основу ее питания. Особенности употребления ими пищи напоминали те, которые характерны для низших животных, — они принюхивались к еде, прежде чем съесть ее, тянулись к пище ртом, точили о кости зубы и т.п. Нет свидетельств о том, чтобы они имели обыкновение прикрывать свое тело или изобретать какую-либо одежду. Возможно, такие дети были сравнительно нечувствительны к жаре и холоду и не испытывали неудобства от того, что ходили обнаженными. Никто не видел, чтобы они плакали, чтобы у них текли слезы или чтобы они смеялись, хотя для них были характерны проявления сильной ярости или нетерпения. Проявления сексуального интереса и соответствующей активности у них либо полностью отсутствовали, либо были чем-то рассеянным, ненаправленным. У этих детей отсутствовало «сознание видовой принадлежности», не было потребности в общении, они избегали людей и часто оказывали предпочтение компании низших животных.
Одним из наиболее важных наблюдений, основанном на изучении диких детей, является то, что связано с развитием пространственного восприятия. В случаях с диким мальчиком Авейрона и Каспаром Хаузером многочисленные трудности в восприятии ими расстояния между предметами, их объемности и плотности были зафиксированы уже во время первоначальных контактов с ними. Итард (49, с. 5) сообщил, что во время первичного обследования дикий мальчик Авейрона был не в состоянии отличать плотные объекты от их изображений. То же было характерно и для Каспара Хаузера. При первом взгляде, брошенном Каспаром Хаузером на пейзаж через окно, он воспринял увиденное как оконный ставень с нанесенным на него цветным рисунком. Такие же трудности он испытывал при оценке размеров объектов, на которые он смотрел с разного расстояния (ср. 72, с. 323).
Некоторые психологи, такие как Деннис (18, 20), склонны были оспаривать сообщения о диких детях. Несомненно то, что в нескольких случаях, проанализированных Зингом, связь с животными, как показывают записи, возникла после достижения детьми возраста установления межличностного общения, или же связь с животными была лишь частичной, и ребенок при этом продолжал поддерживать контакты со взрослыми людьми. Правдой является и то, что данные по некоторым случаям, особенно исторически наиболее ранним, являются весьма скудными, а их авторы — субъектами не очень аккуратными и часто пристрастными, что может быть основанием для сомнений в достоверности этих данных.
Некоторые психологи пошли дальше и предложили другое объяснение для
В своем ответе на критику Денниса Зинг (90) обратил внимание на мозолистые подушечки и другие признаки, приобретенные вследствие продолжительного передвижения на четырех конечностях, а также на степень сноровки, достигнутой в таком способе передвижения, — обстоятельства просто невероятные, если согласиться с тем, что дети до краткого периода изоляции, предшествовавшего их обнаружению, жили в человеческом обществе. Со своей стороны, пищевые предпочтения, наблюдавшиеся во многих подобных случаях, а также физиологические эффекты, являющиеся следствием продолжительного голодания, говорят в пользу длительной изоляции от человеческого общества. Если детей, даже слабоумных, воспитывали люди, то невероятно, чтобы основой их питания являлось сырое мясо. Это особенно справедливо для Индии — именно в ней было найдено много диких детей и именно там у детей не было возможности есть приготовленное мясо (ср. 73).
Зинг также утверждает, что прогресс, достигнутый в обучении детей, возвращенных людям, свидетельствует против гипотезы об их изначальном слабоумии. Однако очевидно, что их достижения вряд ли сопоставимы с уровнем развития нормальных детей соответствующего возраста. Этого вряд ли можно было ожидать не только вследствие длительного периода изоляции, в течение которого у детей не было возможностей для нормального общения с людьми, но также вследствие наличия «негативного переноса» неприемлемых моделей поведения, приобретенных ими во время дикого существования, и которые должны были бы быть забыты, прежде чем достичь какого-либо прогресса. Как исключение, Зинг приводит один случай с индийским «волчьим мальчиком», который действительно был «настоящим идиотом». Этот мальчик так ничему и не смог научиться после возвращения к людям, хотя был уже достаточно взрослым. Сравнительно большой прогресс, достигнутый другими дикими детьми, наводит на мысль о том, что они были изначально нормальными и потеряли человеческие способности только вследствие лишения их в раннем возрасте нужной стимуляции.
Кроме этого, Зинг приводит сообщения надежных свидетелей, показывающих, что по крайней мере двое детей (волчьи дети Миднапура, описанные выше) действительно жили с волками в течение некоторого времени, предшествовавшего их спасению. Этот факт приводится Зингом как ответ на аргумент Денниса об отсутствии прямых свидетельств о воспитании человеческих детей животными. Деннис утверждает, что дети, пойманные вместе с животными, могли оказаться вместе случайно, стремясь спрятаться от преследователей. Деннис настаивает на важности этого пункта для интерпретации поведения диких детей. Он отмечает, что, если ребенок попадает в изоляцию до трех лет, он в состоянии выжить только с помощью животных, если они «принимают» его и заботятся о нем. С другой стороны, если ребенку на момент начала его дикого существования больше трех лет, то у него уже должны быть по крайней мере элементы человеческой речи, локомоции и других подобных функций, в противном случае он является от рождения дефективным.
Интересную гипотезу предложила антрополог Мэриан Смит (73). Поведение диких детей, как считает Смит, можно частично объяснить эмоциональными травмами, произошедшими в результате их изоляции или потери ими своего дома. При этом дети испытывают переживания более сильные, чем при прекращении каких-то занятий. Какая-либо социализация или признаки человеческого поведения, которые были характерны для человека перед его изоляцией, могут перестать существовать вследствие того, что в последующем опыте ребенка они не подкрепляются. Чем дольше продолжается период дикого существования ребенка по сравнению с более ранним периодом человеческого воспитания, тем труднее заметить признаки социализации. Данная гипотеза помогает понять, почему такие формы социального поведения, как соответствующая походка и речь, развивавшиеся у ребенка до его изоляции, не были зафиксированы у диких детей.
Возражения, выдвинутые Деннисом и другими учеными, должны учитываться при оценке сообщений об обнаружении детей, которых объявляют дикими. Вероятно, что эти возражения применимы к большинству случаев, обсуждавшихся Зингом и другими исследователями. С другой стороны, существуют достоверные свидетельства о том, что три или четыре случая действительно представляют собой примеры продолжительной изоляции детей от общества. Дикий мальчик Авейрона, волчьи дети Миднапура, Каспар Хаузер и, возможно, еще один или два случая относятся к случаям данной категории. Изучение этих случаев помогает пролить свет на некоторые факты человеческого развития, установленные при помощи других, лучше контролируемых исследований.
Культурные различия в традициях воспитания детей. Другим видом «естественного эксперимента» можно считать
Тестирование десяти таких младенцев в возрасте от 4 месяцев до 1 года (ср. 14) показало существенное замедление развития их поведения. Мало кто из них был способен при представившейся возможности на спонтанную реакцию. У них была плохая координация, а хватательные движения появлялись с запозданием. Только один из десяти младенцев был в состоянии ползать ранее, чем в 1 год, хотя все они могли сидеть без поддержки. С другой стороны, их социальные реакции были достаточно развиты, видимо, вследствие того что семьи были большими и их члены были единственными стимуляторами в окружающей младенца среде. У детей старше одного года были нормальные социальные реакции, способности к обучению и «интеллектуальная продуктивность», но они страдали от недостаточно развитой координации и неспособности активно выражать свои желания. Увидев незнакомые предметы, дети проявляли интерес и желание рассмотреть их, но делали это робко и неуклюже, в новых для себя ситуациях им часто требовалась помощь взрослых.
Обычаи, ограничивающие двигательную активность младенцев, были также зафиксированы у некоторых индейских племен Америки, таких как Наваго и Гопи. В этих племенах новорожденные дети туго пеленались в одеяло и тщательно привязывались к жесткой доске. Находясь в таком положении, младенец не мог поворачиваться и даже двигать руками и ногами. В первые три месяца своей жизни он находился в этих пеленках, его чистили и купали каждый день, лишь в течение часа. Позже ему позволяли больше времени проводить вне колыбели. Деннис (17) сообщает, что, несмотря на такие жесткие ограничения движений, когда дети Гопи и Наваго освобождаются, они могут так же хорошо сидеть, ползать и ходить, как и белые американские дети. Во время коротких периодов, когда их освобождают от пеленок, они обычным образом выгибаются, тянутся к предметам и тащат их в рот, берут игрушки и засовывают их в рот и демонстрируют другие признаки характерного моторного поведения ребенка с неограничиваемой активностью. Представляет интерес также то обстоятельство, что исследователи не нашли существенной разницы между средним возрастом, когда начинали ходить дети, воспитывавшиеся в такой традиционной манере, и средним возрастом, когда начинали ходить обычные белые американские дети. В группе из 63 детей, воспитывавшихся согласно обычаям Гопи, средний возраст, когда они начинали ходить, был 14,95 месяцев; а для 42 детей Гопи, воспитывавшихся без жесткого ограничения движений, этот возраст составил 15,05 месяцев. Такое различие не является статистически существенным (22).
Аналогичные результаты были получены в ходе исследования 110 японских и корейских детей, протестированных в возрасте между 1 месяцем и 3 годами (46). По результатам тестов на моторное развитие эти дети не показали отставания, хотя согласно традициям матери носили их на своих спинах ежедневно в течение длительного времени.
Результаты этих исследований не вполне соответствуют друг другу, первое исследование показало гораздо большие функциональные нарушения, явившиеся следствием недостатка двигательной активности, чем было выявлено в ходе двух других исследований. Конкретные данные, очевидно, зависят от множества факторов, таких как природа и степень ограничений, а также возраст, в котором они прекращаются. Весьма вероятно, например, что сами по себе моторные ограничения приводят к гораздо меньшим нарушениям функциональной деятельности, чем в сочетании с недостаточностью сенсорной стимуляции и социальных контактов. На развитие конкретных функций, находившихся под наблюдением, воспитание, несомненно, влияло по-разному. Исследователи в целом сходятся во мнении, что определенные простые моторные функции, появляющиеся в раннем младенчестве, почти полностью зависят от созревания, в то время как многие другие функции для своего нормального развития требуют сравнительно коротких периодов для наработки.
Многие современные исследования культурных различий в практике воспитания детей касались воздействия определенных аспектов занятий с ребенком на его последующее
Можно упомянуть о наличии большого массива доступных данных по влиянию
Подготовка и индивидуальные различия. Еще один тип исследований, касающихся влияния предшествовавшего опыта на поведенческое развитие, в котором рассматриваются обучение и воспитание в школе, будет обсуждаться нами в главе 7. Эти исследования включают в себя множество контролируемых лабораторных экспериментов, помогающих ответить на вопрос, увеличиваются или уменьшаются индивидуальные различия после периода единообразного развития некоторой простой функции.
Испытуемыми в таких исследованиях обычно являются студенты колледжа или его выпускники, иногда ученики школ.
Впечатляющий по своим размерам объем данных накоплен по проблеме воздействия школьного образования. К этой категории относится группа противоречивых исследований, касающихся воздействия детского сада на IQ ребенка. Несколько исследователей собрали данные об эффективности специальных подготовительных программ в развитии интеллекта слабоумных испытуемых. Это исследование будет обсуждаться в главе 12, посвященной проблеме интеллектуальной неполноценности. Еще одна группа исследований связана с отношениями между школьным обучением и умственным развитием в старшем возрасте. Далее следует анализ долговременных тенденций в развитии общего интеллектуального уровня населения по отношению к периодически происходящим изменениям в сфере образования.
Пятый и последний подход к анализу наследственности и окружающей среды заключается в статистическом анализе родовых сходств и различий. Этот подход будет обсуждаться в главе 9. Интерпретация таких сходств и различий осложняется тем, что близкие родственники, как правило, живут вместе. Окружающая среда для людей, живущих в одном и том же доме, является более сходной, чем в какой-либо другой ситуации, внешней по отношению к экспериментальной ситуации. В результате два класса факторов — наследственных и связанных с окружающей средой — действуют в одном и том же направлении, а именно, увеличивают сходство внутри обыкновенной семьи, в отличие от влияния на случайно выбранных индивидов. Чем теснее наследственное отношение, тем сильнее проявляется близость окружающей среды. Так, родители и дети, братья и сестры обычно живут в одном и том же доме; в то время как более далекие родственники, такие как дяди и племянники или двоюродные братья и сестры, вступают друг с другом в более редкие контакты. Вследствие сходства жилищных условий на родственников воздействует не только общая окружающая среда, но и они сами, образуя окружающую среду друг для друга, через такое взаимное воздействие могут становиться в некоторых отношениях еще более близкими. Так или иначе, семьи представляют собой отличный пример воздействия окружающей среды, усиливающей поведенческое сходство их членов. В то же время тот факт, что члены одной и той же семьи несут в себе в разной степени общую наследственность, делает родовые сходства главным источником данных для проверки гипотез, связанных с факторами наследственности. При этом следует помнить о том, что само по себе родовое сходство не может рассматриваться как свидетельство наличия общей наследственности. Чтобы продемонстрировать действие наследственных факторов, потребуются дополнительные анализы и контрольные процедуры.
Исследования родословной. Проявление семейных родословных относительно некоей особой и легко идентифицируемой характеристики может способствовать получению ценных данных, касающихся факторов наследственности. Методы анализа родословной, используемые генетиками, особенно хорошо зарекомендовали себя при исследованиях простых физических отклонений, таких как альбиносность, перепонки на пальцах, наличие лишних пальцев или «копыта» и множества других более редких и более серьезных уродств или патологий. При помощи этих техник можно также анализировать некоторые психологические отклонения.
Однако применение анализа родословной к большинству психологических качеств обычно сталкивается с непреодолимыми препятствиями. Главная трудность связана со множественностью факторов, влияющих на психологические свойства. В результате большинство исследований родословной в психологии ограничиваются демонстрацией того, что определенные качества, такие как слабоумие, безумие, музыкальный талант или выдающиеся научные способности, имеют тенденцию к воспроизводству в роду. Эти данные — их мы будем рассматривать в главе 9 — являются по существу описательными и практически не могут служить основанием для объяснения наблюдаемых сходств.
Корреляционные исследования родовых сходств. Степень сходства между родственниками можно выразить с помощью корреляционного коэффициента (ср. гл. 1). Например, высокая положительная корреляция между результатами теста на умственные способности отцов и сыновей будет показывать то, что сыновья, отцы которых обладают высокими показателями, имеют тенденцию демонстрировать такие же высокие результаты. Большинство корреляционных исследований касалось степени родства между показателями родителей и детей, между близнецами и другими детьми данных родителей. Меньшая часть исследований имела дело с более слабыми родственными отношениями, такими как между двоюродными братьями и сестрами или между дедами и внуками.
Во всех исследованиях, связанных с близнецами, важным является различение однояйцевых и разнояйцевых близнецов (ср. гл. 3). Напомним, что однояйцевые близнецы несут в себе одну и ту же наследственность. Поэтому все различия между ними являются результатом пренатального или постнатального воздействия факторов окружающей среды. С другой стороны, разнояйцевые близнецы имеют большее генетическое сходство, чем простые братья и сестры. Таким образом, сравнивание степени сходства психологических качеств между однояйцевыми близнецами, разнояйцевыми близнецами и просто братьями и сестрами открывает новые возможности для анализа действия наследственности и окружающей среды.
Исследования особых родовых отношений. Некоторые исследователи увидели в исследовании приемных детей новые возможности для определения влияния наследственности и окружающей среды. Такие исследования позволяют получить несколько групп данных. Например, можно изучать те изменения, которые произошли в результате изменения места жительства ребенка после переезда в дом приемных родителей, а также степень сходства приемных детей со своими собственными родителями, с приемными родителями и с детьми приемных родителей. Этот метод может применяться также для исследования однояйцевых близнецов, воспитывавшихся отдельно друг от друга у разных приемных родителей[17]. С этой точки зрения также могут быть исследованы дети, воспитывавшиеся в интернатах. Основные результаты исследований, проведенных в русле каждого из перечисленных подходов, будут даны в главе 9.
Рассматривая методы, которыми пользуются психологи для исследования наследственности и окружающей среды, важно помнить о той проблеме, о которой шла речь в 3 главе. Это вопрос не о том, от чего зависит данный признак — от фактора наследственности или от факторов окружающей среды — и не о степени влияния каждого из факторов. Поднимаемый данными исследованиями вопрос заключается в идентификации специфических наследственных факторов и факторов окружающей среды, влияющих на развитие каждой функции, и состоит в том, как эти факторы связаны с возникновением наблюдаемых индивидуальных различий. Самое непосредственное отношение к этим проблемам имеет исследование влияния «структурных», или органических, условий на развитие поведения, поскольку наследственность может влиять на поведение только через соответствующие структурные характеристики.
Классификация методов, используемых в психологических исследованиях влияния факторов наследственности и окружающей среды на поведение, может включать пять основных групп: селективное выведение, исследования нормативов развивающегося поведения, исследование структурных факторов в развитии поведения, исследование влияния на поведение предшествующего опыта и статистический анализ родовых сходств и различий.
Исследования роли
Среди исследований
Пятым общим методом является статистический анализ родовых сходств и различий, подробнее он будет рассмотрен в главе 9. Сюда относятся исследования родословных или историй родов; всевозможные родственные корреляции, выражаемые в показателях тестов; исследования воспитывавшихся порознь однояйцевых близнецов, приемных детей и детей, выросших в условиях интерната.
1. Ames, Louise В. The sequential patterning of prone progression in the
human infant.
2. Beach, F. A., and Jaynes, J. Effects of early experience upon the behavior
of animals.
3. Brudy, E. G. Genetic basis of spontaneous activity in the albino rat.
4. Carmichael. L. The development of behavior in vertebrates experimentally
removed from the influence of external stimulation.
5. Carmichael, L. A further study of the development of behavior in vertebrates
experimentally removed from the influence of external stimulation.
6. Carmichael, L. A further experimental study of the development of behavior.
7. Carmichael, L. An experimental study in the prenatal guinea pig of the origin
and development of reflexes and patterns of behavior in relation to the stimulation of specific receptor areas during the period of active fetal life.
8. Carmichael, L. Ontogenetic development. In S. S. Stevens (Ed.),
9. Carmichael, L. The onset and early development of behavior. In L. Car-
michael (Ed.),
10. Coghill, С. Е.
Press, 1929.
11. Coronios, J. D. Development of behavior in the fetal cat.
12. Cruikshank, Ruth M. Animal infancy. In L. Carmichael (Ed.),
13. Craze, W. W. Maturation and learning in chicks. /.
19, 371–409.
14. Danzinger, L., and Frankl, L. Zum Problem der Funktionsreifung. Erster
Berieht uber Entwickhlugsprufungen en albanischen Kindern. Z
15. Davis, A., and Havighurst, R. J. Social class and color differences in child
rearing.
16. Dennis, W. Infant development under conditions of restricted practice
and of minimum social stimulation: a preliminary report. /.
17. Dennis, W. Does culture appreciably affect patterns of infant behavior?
18. Dennis, W. The significance of feral man.
425-432.
19. Dennis, W. Is the newborn infant's repertoire learned or instinctive?
20. Dennis, W. A further analysis of reports of wild children.
1951, 22, 153–158.
21. Dennis, W. and Dennis, Marsena, G. Behavioral development in the first year
as shown hv forty biographies.
22. Dennis, W., and Dennis, Marsena G. The effect of cradling practices
upon the onset of walking in Hopi children. /.
23. Forgus, R. H. The effect of early perceptual learning on the behavioral
organization of adult rats. /.
24. Fromme, A. An experimental study of the factors of maturation and practice
in the behavioral development of the embryo of the frog,
25. Fuller, J. I. and Scott, J. P. Heredity and learning ability in infrahuman
mammals.
26. Furchtgott, E. Behavioral effects of ionizing radiations.
1956, 53, 320–334.
27. Gesell, A.
28. Gesell, A, The ontogenesis of infant behavior. In L. Carmichael (Ed.),
29. Gesell, A., et al.
30. Gesell, A. and Amatruda, Catherine S.
N.Y.: Hoeber, 1917.
31. Cesell. A., and Thompson, Helen. Twins T and С from infancy to adoles-
cence: a biogenetic study of individual differences by the method of co-twin control.
32. Gos, M. Les reflexes conditionnels chez I'embrvon d'oiseau.
33. Hall, С S. Emotionality in the rat.V.
1936, 22, 61–68, 345–352; 1937, 24, 369–376.
34. Hall, C. S. The genetics of behavior. In S. S. Stevens (Ed.),
35. Hall, C. S., and Klein, S. J. Individual differences in aggressiveness in rats.
36. Halverson, H. M. The acquisition of skill in infancy.
1933, 43, 3-48.
37. Harris, H. E. An analysis of the maze-learning scores of bright and dull rats
with reference to motivational factors.
38. Hayes, Catherine. The
39. Hayes, K. J., and Hayes, Catherine. The intellectual development of a
home-raised chimpanzee.
40. Hayes, K. J., and Hayes, Catherine. Imitation in a home-raised chimpanzee.
41. Hayes, K. J., and Hayes, Catherine. The cultural capacity of the chimpanzee
42. Hebb, D. O.
43. Heron, W. T. The inheritance of brightness and dullness in maze learning
ability in the rat. /.
44. Hilgard, J. K. The effect of early and delayed practice of memory and
motor performances studied by the method of co-twin control.
45. Hirsch, J., and Tryon, R. C. Mass screening and reliable individual
measurement in the experimental behavior genetics of lower organisms.
46. Hofstaetter, P. R. Testuntersuchungen an japanischcn Kindern und das
Reifungsproblem.
47. Hooker, D.
1952.
48. Hunt, E. L. Establishment of conditioned responses in chick embryos. /.
49. Hard, 1. M. C.
N.Y.: Appleton-Century-Crofts, 1932.
50. Kellogg, W. N. A further note on the «wolf children» of India.
51. Kellogg, W. N., and Kellogg, Luella A.
House. McGraw-Hill, 1933.
52. Kuo, Z. Y. Ontogeny of embryonic behavior in aves: I. The chronology and
general nature of the behavior of the chick embryo. /.
53. Kuo, Z. Y. Ontogeny of embryonic behavior in aves: III. The structure and
environmental factors in embryonic behavior. /.
54. Kuo, Z. Y. Ontogeny of embryonic behavior in aves: IV. The influence of
embryonic movements upon the behavior after hatching. /.
55. Kuo, Z. Y Development of acetylcholine in the chick embryo. /.
1939, 2, 488–493.
56. Levinson, B. Effects of fetal radiation on learning. /.
1952, 45, 140–145.
57. Lindsley, D. В. Heart and brain potentials of human fetuses in utero.
58. Luchins, A. S., und Korgus, К. Н. The effect of differential post-weaning
environment on the rigidity of an animal's behavior.
59. Milner, Esther. A study of the relationships between reading readiness in
grade one school children and patterns of parent-child interaction.
60. Munn, N. L.
Mifflin, 1955.
61. Pratt, К. С The neonate. In L. Cannichael (Ed.),
(2nd Ed.) N.Y.: Wiley, 1954, 215–291.
62. Riess, B. F. The effect of altered environment and of age on mother
young relationships among animals. Ann. N. Y.
63. Rundqist, E. A. Inheritance of spontaneous activity in rats.
1933, 16, 415–438.
64. Scott, J. P. Genetic differences in the social behavior of inbred strains of
mice. /.
65. Scott, J. P. New directions in the genetic study of personality and intelligence.
66. Scott, J. P., and Charles, Margaret S. Some problems of heredity and
social behavior. /.
67. Scott, J. P., and Fuller, J. L. Research on genetics and social behavior at
the Roscoe B. Jackson Memorial Laboratory, 1946—1951-a progress report.
68. Searle, L. V. The organization of hereditary maze-brightness and
mazedullness.
69. Shirley, Mary M. A motor sequence favors the maturation theory.
70. Shirley, Mary M.
development. Vol. II. Intellectual development. Minneapolis: Univer. Minn. Press, 1931, 1933.
71. Silverman, W. Shapiro. F., and Heron. W. T. Brain weight and maze learning
in rats. /.
72. Singh. J. A. L., and Zingg, R. M.
1942.
73. Smith, Marian W. Wild children and the principle of reinforcement.
74. Spelt, D. K. The conditioning of the human fetus in utero
1948, 38, 338–346.
75. Sperry, R. W. Mechanisms of neural maturation. In S. S. Stevens
(Ed.),
76. Squres, P. С. Wolf children of India.
315.
77. Stockard, C. H., Anderson, O. D., and James, W. T.
78. Stone, C. P. Methodological resources for the experimental study of innate
behavior as related to environmental factors.
79. Stratton, С М. Jungle children.
80. Straver, L. C. Language and growth: the relative efficacy of early and
deferred vocabulary training, studied by the method of co-twin control,
81. Tait, С D., Jr., Wall, P. D., Balmuth, Muriel, and Kaplan, S. J. Behavioral
changes following radiation. II. Maternal behavior and maze performance. USAF,
82. Thompson, W. R., and Heron, W. The effects of restricting early experience
on the problem-solving capacity of dogs.
83. Thompson, W. R. The heredity-environment problem,
84. Tinbergen, N.
85. Tryon, R. C. The genetics of learning ability in rats: preliminary report.
Univer.
86. Tryon, R. C. Genetic differences in maze-learning ability in rats. 39th
Year*.,
87. Whiting, J. W. M., and Child, I. L.
88. Wickens, D. D., and Wickens, Carol. A study of conditioning in the
neonate. /.
89. Zingg, R. M. Feral man and extreme cases of isolation.
1940, 53, 487–517.
90. Zingg, R. M. Reply to Professor Dennis' «The significance of feral man».
| © 2024 Библиотека RealLib.org (support [a t] reallib.org) |