"Основы физиологии высшей нервной деятельности" - читать интересную книгу автора (Коган Александр Борисович)
Глава 6 СОН И СНОВИДЕНИЯ
Периодическая смена деятельности и покоя наблюдается у всех живых существ. Однако под сном принято понимать совершенно определенное явление.
Сон рыб. Сон рыб зависит от их экологии. Так, ночные хищники спят большую часть дня и охотятся ночью, однако сон их своеобразен. Например, карликовый сомик и ночью, временами впадая в «сонное» состояние, остается неподвижным, сердечные сокращения замедляются, но тонус мускулатуры — напряженный. Это состояние обозначили как «ночной покой» в отличие от «дневного покоя». Однако по показателям частоты сердечных сокращений дневной покой этого ночного хищника оказался более глубоким, чем ночной (соответственно 26 и 36 сокращений сердца/мин при 48 сокращениях при бодрствовании). Дыхание также становилось гораздо реже при дневном покое (15 вдохов/мин), чем при ночном покое (34 вдохов/мин), при гораздо более частом дыхании в состоянии бодрствования (68 вдохов/мин). Учет сравнительной длительности бодрствования и покоя карликового сомика при круглосуточной естественной освещенности показал сходные результаты (рис. 38), что свидетельствует о наличии у него внутреннего ритма смены бодрствования и покоя.
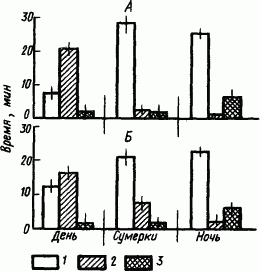 |
Рис. 38. Средняя длительность бодрствования
Другой характер суточной периодики поведения наблюдали у рыб, питающихся в дневное время. Например, у кефали урежение сердечных сокращений во время ночного покоя более выражено, чем во время дневного. Однако смена покоя и бодрствования не сопровождается у рыб характерными изменениями электрической активности мозга, которые проявляются у птиц и млекопитающих и свидетельствуют о развитии в высших отделах нервной системы определенных стадий процессов сонного торможения. Поэтому такие состояния покоя у низших позвоночных предшествующие в эволюции сну высших животных, получили название
Сон амфибий. У амфибий суточное распределение бодрствования и покоя нерегулярно и имеет резко выраженные экологические различия. Так, у травяной лягушки, ведущей ночной образ жизни, днем преобладает состояние покоя с урежением сердечных сокращений. У озерной лягушки, подстерегающей добычу 80–90 % времени суток занимает состояние неподвижности, которое внешне является покоем, но без потери мышечного тонуса и урежения сердечных сокращений, чем отличается от истинного сноподобного покоя. Вместе с тем у амфибий можно наблюдать периоды более глубокого торможения, которые сменяются фазическими движениями, сопровождающимися быстрыми колебаниями электрических потенциалов в крыше среднего и отделах переднего мозга.
Сон рептилий. У рептилий суточная периодичность смены бодрствования и покоя становится более четкой. Например, болотная черепаха днем бодрствует или находится в состоянии дневного покоя (рис. 39). Однако дневной покой черепахи выражается лишь в неподвижности при открытых глазах, сохранении мышечного тонуса и способности к ориентировочным реакциям. Зато ночной покой характеризуется углублением состояния общего торможения, расслаблением мускулатуры при закрытых глазах. При этом в структурах головного мозга появляются медленные колебания потенциалов в диапазоне 3–7 Гц, сходные с «сонными потенциалами» высших животных.
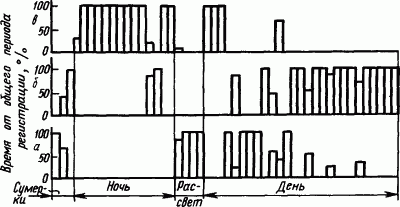 |
Рис. 39. Суточная периодика бодрствования
Сон птиц. Птицам свойственна четкая определенность суточной периодики смены сна и бодрствования, зависящей от условий их жизни. Так, у типичных дневных птиц — кур — бодрствование днем занимает 33 % времени суток, хотя и перемежается периодами дневного покоя (18 % суточного времени). При этом курица неподвижна, с открытыми глазами, сохраняет мышечный тонус и может реагировать ориентировочной реакцией на слабые раздражители. Ночной покой кур, по наблюдениям, длится 49 % времени суток. В это время куры с закрытыми глазами, расслабив мускулатуру, погружаются в состояние общего торможения. По показателям поведения, вегетативных реакций и электрических потенциалов мозга это истинный сон в том виде, в каком он проявляется у высших животных.
У птиц впервые четко обнаружено по показателям электрической активности мозга деление ночного сна на так называемую
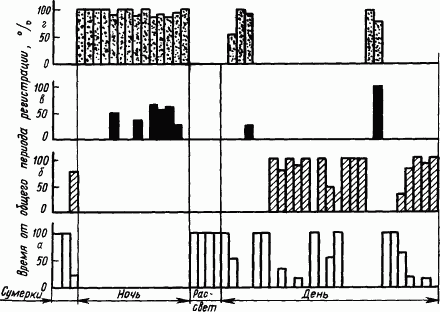 |
Рис. 40. Суточная периодика бодрствования
Типичные ночные птицы — совы — спят днем и бодрствуют ночью. По некоторым наблюдениям, время их бодрствования составляет 47 % общего времени суток. В дневном сне МС занимает 25 %, а ПС —3 % времени суток. Однако кроме активного сна у сов наблюдаются и периоды дневного покоя, составляющие 25 % времени суток. По своим проявлениям дневной покой совы, застывшей с открытыми глазами, напоминает каталептическое состояние (в это время экспериментатор мог брать сову за клюв и придавать ее голове неестественное положение, которое длительно сохранялось).
Сон млекопитающих. Эволюция приспособительного поведения животных привела на уровне теплокровных к регулярному чередованию бодрствования и покоя, принявшего форму глубокого сонного торможения. При этом происходило усложнение структуры сна у разных животных в соответствии с видовыми особенностями их образа жизни. Четко разграничились МС и ПС сна. Существуют данные о том, что у дельфинов медленные дельта-волны могут возникать лишь в каком-либо одном полушарии, а в другом сохраняются альфа-подобные волны, свойственные состоянию бодрствования. Это позволило предположить, что дельфины «спят одним из полушарий». Высокой степени тонкого приспособления достигла организация смены сна и бодрствования у хищных в связи с их охотой. Общеизвестна способность кошки засыпать и просыпаться многократно на протяжении сравнительно небольшого отрезка времени.
Сон человека. Сон человека — это состояние избирательного торможения процессов высшей нервной деятельности, которое возникало как защитная неподвижность в ночное время, сформировало нейрогуморальные механизмы регулирования смены сна и бодрствования и подчинило своему суточному ритму все функции организма вплоть до обмена веществ. Время сна дифференцировано на МС и ПС главным образом для пластических репарационных процессов, обработки накопившейся информации и консолидации долговременной памяти.
Суточный ритм жизни человека сложился из активной деятельности днем и глубокого сна ночью. Современный человек спит в среднем 7–8 ч/сут. Однако с возрастом продолжительность сна меняется. Так, новорожденный ребенок, кроме времени кормления, спит непрерывно. В возрасте 3–5 месяцев он спит уже только 17–18 ч, а к 2–3 годам жизни время сна уменьшается примерно до 12 ч/сут. Нормальная для взрослого человека продолжительность сна 7–8 ч/сут устанавливается к 15–18 годам жизни. В старческом возрасте обычно спят меньше. Вместе с тем длительность сна может быть различна у разных людей. Из жизнеописания Петра Первого следует, что он спал не более 5–6 ч, и этого было ему достаточно. Описаны также многочисленные случаи, когда человек довольствовался еще более ограниченным временем сна.
С наступлением сна расслабляется вся мускулатура. Однако это происходит в известной последовательности. Когда человека одолевает сон, он начинает, как говорят, «клевать носом», так как мышцы шеи, поддерживающие голову, постепенно теряют свой тонус. Затем расслабляются мышцы, сохраняющие позу и положение тела. Поэтому человек спит лежа, когда все его мышцы максимально расслаблены. При этом тормозятся и некоторые вегетативные функции: замедляются сердечные сокращения, снижается артериальное кровяное давление, становится более редким дыхание, уменьшаются траты энергии в организме. В то же время усиливается секреция желудочных и кишечных желез, более интенсивно идет всасывание, активируются многие синтетические процессы.
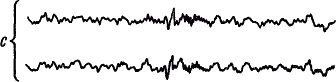
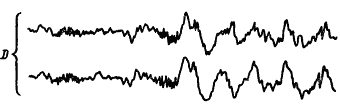
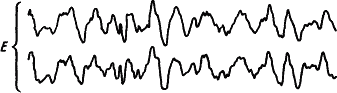
Динамику электрической активности мозга в процессе развития и течения сна у человека изучали многие исследователи. Была предложена классификация стадий сна на основе изменения уровня сознания и формы электроэнцефалограммы (А. Лумис и др., 1937). К описанным стадиям затем добавили стадию «быстрых движений глаз», названную стадией REM (от англ. rapid eye movement) (В. Демент, Н. Клайтман, 1957) и более известную под названием парадоксальной стадии сна (М. Жуве, 1967). На рис. 41 показаны основные стадии развития естественного сна у человека.
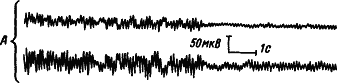 |
 |
 |
 |
 |
 |
Рис. 41. Изменения электроэнцефалограммы человека при развитии сна, объяснения см. в тексте (по А.Н. Шеповальникову)
Стадия
Стадии
У человека также проявляются избирательность порога пробуждения, наличие своеобразных «сторожевых пунктов». Так, мать у постели больного ребенка после бессонных ночей засыпает так крепко, что ее не разбудит ни громкий разговор, ни хлопанье дверью, ни бравурная музыка по радио. Но она мгновенно просыпается, услышав тихий стон ребенка.
В некоторые стадии сна (главным образом, ПС) могут возникать, хотя и в неадекватной, искаженной форме, образы внешнего мира в виде
В сновидениях отражаются прошлая жизнь человека и его переживания (З. Фрейд, 1933). Анализ снов 21 студента, разбуженных в период ПС, показал, что содержание почти всех сновидений основывалось на пережитом (Дж. Матсумото, 1985).
Другим источником сновидений могут быть текущие ощущения. Например, человеку приснилось, что он пошел в дальний поход, переходил замерзшую реку, провалился под лед и ноги оказались в ледяной воде. В действительности у него с ног сползло одеяло. В этом и других случаях проявляется удивительное свойство сновидений за кратчайший срок пронести в сонном сознании человека фантастические события, развивающиеся в течение длительного времени. Об этом очень наглядно свидетельствует пример, приведенный одним французским исследователем, который спал в постели под пологом, подвешенным на палке. Ему приснился длинный сон о том, как его схватили, судили, приговорили к смерти, привели к гильотине, положили голову на плаху, нож гильотины упал на шею... и он проснулся, разбуженный ударом по шее сорвавшейся палкой полога. За мгновенье, прошедшее от удара до пробуждения, в его мыслях пронеслись все события этого сновидения.
О связи сновидений с бывшими впечатлениями убедительно свидетельствует тот факт, что в своих снах слепорожденные не видят зрительных сцен, а глухорожденные не слышат голосов и звуков.
Все показатели поведения и состояния большинства других функций организма указывают на то, что сон представляет собой в нейрофизиологическом понимании разлитое торможение процессов высшей нервной деятельности, особенно выраженное в коре больших полушарий головного мозга. Условиями его развития могут быть торможение в результате ограничения афферентации и условное торможение на время суток («пассивный» и «активный» сон, по И.П. Павлову, 1932), охранительное торможение, вызываемое утомлением корковых клеток, торможение под действием специфических медиаторов, безусловно-рефлекторное торможение в процессе инстинктивного поведения.
О последовательности охвата торможением мозговых структур можно судить по тому, как при засыпании сначала теряется ясность мышления, затем исчезает сознание, потом расслабляется мускулатура и наконец снижается деятельность вегетативных органов. Если рассматривать развитие и распространение процессов торможения по электрическим проявлениям медленноволновой стадии сна — «сонным потенциалам» (дельта-волны), то, как видно из рис. 42, при естественном засыпании кошки торможение возникает вначале в лобных областях коры, потом распространяется на ее теменные и затылочные области, а затем спускается в подкорковые и стволовые структуры мозга. По-видимому, ведущая роль лобных областей коры больших полушарий у кошки в развитии «сонных потенциалов», как и в генерации фоновой ритмики, определяется тем, что они имеют более тесные связи с восходящими неспецифическими системами мозга. Однако у человека, мозг которого организован так, что его фоновая ритмика наиболее выражена в затылочных областях коры, развитие «сонных потенциалов» начинается именно из них.
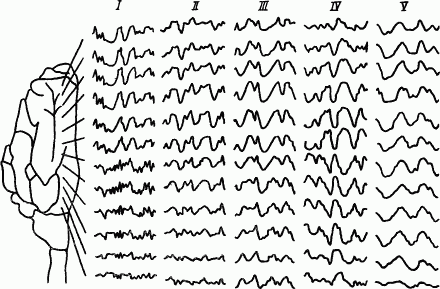 |
Рис. 42. Распространение дельта-волн по поверхности коры мозга кошки при развитии сна (по А.Б. Когану):
Послойный анализ электрической активности коры мозга кролика показал, что источником дельта-волн во время наркотического сна являются тела пирамидных нейронов
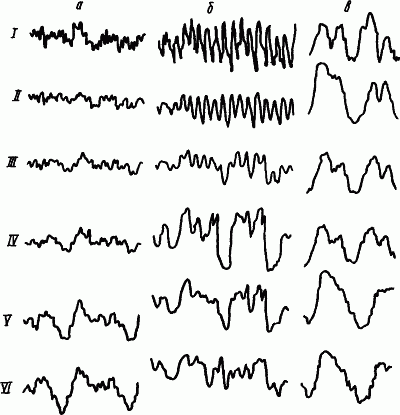 |
Рис. 43. Распространение дельта-волн по слоям коры мозга кошки
Уже на стадии сонливости
Нейрофизиологический механизм сна постепенно раскрывался по мере накопления фактов о естественных и искусственных условиях его возникновения. Сведения о том, что утомленный человек быстрее засыпает и крепче спит, указывали на возможную связь деятельности мозга с состоянием гуморальных систем. Патологические изменения нервной ткани в области начала сильвиева водопровода при «сонной болезни» — летаргическом энцефалите (И. Маутнер, 1820) показали, что центры мозгового ствола могут иметь отношение к явлениям сна. В лаборатории создателя учения об условных рефлексах — И.П. Павлова сон вызывали у собак сигналами условного торможения, развивающегося в высших отделах мозга.
Вместе с тем оказалось, что раздражением некоторых структур промежуточного мозга у кошек можно вызвать полную картину естественного сна. В экспериментах с перерезками ствола мозга на разном уровне наблюдали сохранение электрокортикограммы, характерной для бодрствования, если разрез был ниже продолговатого мозга, и развитие «сонных потенциалов», когда он был выше среднего мозга. Наконец, прямым раздражением ретикулярной формации среднего мозга было показано ее активирующее действие на высшие отделы мозга (Д. Моруцци, Г. Мэгун, 1949). Прекращение этого действия погружает животное в сон, а его возобновление вызывает «реакцию пробуждения». Наряду с активирующей системой, десинхронизирующей электрокортикограмму, были обнаружены стволовые гипногенные системы, которые при низкочастотном раздражении синхронизируют электрокортикограмму и приводят к состоянию сна. К ним относятся базальные отделы переднего мозга, медиальные структуры таламуса, некоторые структуры хвостатого ядра продолговатого мозга, гиппокампа и гипоталамуса. Раздражение дорсомедиальных отделов варолиева моста вызывает возникновение парадоксальной стадии сна, а повреждение ядер шва создает бессонницу.
Исследование процессов сонного торможения на нейронном уровне показало, что при этом не происходит общего затормаживания всех корковых нейронов. Часть нейронов действительно замедляет свою фоновую импульсацию, но другая часть — учащает. Такие неоднозначные изменения частоты импульсов нейронов при развитии сна наблюдали также в гиппокампе, таламусе и других подкорковых и стволовых структурах мозга. Отсюда следует, что сонное торможение создается определенной организацией возбуждающихся и тормозящихся нейронов. Эта организация обнаружила следующие характерные особенности поведения корковых нейронов в разные стадии сна.
1. При подсчете средней частоты фоновой импульсации всех нейронов, как замедляющих, так и учащающих свой разряд, выяснили, что
2. Перестраивается структура импульсных потоков.
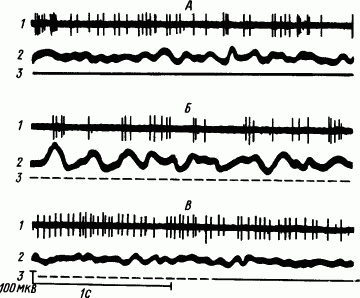 |
Рис. 44. Фоновая импульсация нейронов теменной коры мозга кошки в состоянии бодрствования
Таблица 8. Распределение различной структуры импульсных потоков теменной коры кошки в разные стадии сна, %
 |
Во время МС каждая пачка объединяет 5–15 импульсов с интервалами молчания между пачками 200–500 мс. Такое формирование структуры импульсного потока характерно для влияний восходящей неспецифической таламокортикальной системы, приводящих к развитию тормозных процессов в коре. Во время ПС большинство нейронов возвращается к непрерывно-аритмическому типу фоновой импульсации. Однако часть нейронов сохраняет ее пачечно-групповую структуру, т.е. в этой стадии возникает некоторое разделение возбуждающих и тормозящих механизмов корковой деятельности.
3. Исследование взаимосвязи импульсации корковых нейронов ползало, что в
Таблица 9. Взаимосвязь импульсных потоков (по коэффициентам корреляции) разноудаленных нейронов теменной коры мозга кошки в разные стадии сна
 |
Такая синхронность разрядов обширных популяций нейронов, видимо, также обусловлена диффузными влияниями восходящей таламокортикальной системы, организующей развитие процессов сонного торможения. Этим объясняется и то, что возникающие в МС пачки импульсов оказываются приуроченными к фазам дельта-волн. Аналогичные изменения корреляции импульсные разрядов корковых нейронов наблюдались при раздражении неспецифических структур таламуса, приводящем к возникновению веретен. При переходе к ПС таламические влияния ослабевают и усиливается активность структур среднего мозга.
4. При испытании реактивности нейронов теменной коры кошки оказалось, что
Таблица 10. Динамика реактивности корковых нейронов в процессе развития сна
 |
Существует некоторое различие в динамике ответов на афферентные раздражения и прямую стимуляцию входных в кору путей, в частности практически неизменный высокий процент заторможенных нейронов в одном случае и сравнительно малое число нереагирующих — в другом. Эти различия могут указывать на то, что развитие сна затрагивает и специфические релейные ядра таламуса, затрудняя прохождение через них афферентных сигналов. Нейроны, реагировавшие возбуждением при бодрствовании животного, могли при развитии сна не только перестать возбуждаться, но и начинали реагировать торможением. Примечательно, что в стадии МС уменьшается вариабельность импульсных реакций нейронов на раздражения, что можно объяснить как результат резкого ограничения полисенсорного реагирования.
5. Построение гистограмм распределения величин латентных периодов импульсных ответов раздельно для их значений менее и более 5 мс выявило, что
6. Возникающий в ответ на раздражение сложный узор мозаики активности элементарных нейронных ансамблей с наступлением сна претерпевает глубокие изменения.
Это наглядно показывает математическая реконструкция мозаик активности нейронов, построенная по данным одновременной мультиэлектродной регистрации нейронных реакций методом поэтапной оптимизации. При этом во время МС микроочаги возбуждающихся нейронов, ранее разделенные тормозящимися, сливаются между собой, образуя массивные зоны возбуждения наряду с массивами тормозящихся нейронов. В период ПС эти массивы дробятся, но не достигают степени дробности, характерной для состояния бодрствования.
7. Если считать, что нейроны передают информацию потоками импульсов, то можно попытаться оценить их информационную деятельность в понятиях теории информации, разработанной для технических систем связи (К. Шеннон, 1963). Результаты такой оценки представлены в табл. 11.
Таблица 11. Информационные характеристики импульсной активности нейронов теменной коры мозга кошки в разные фазы сна
 |
Все перечисленные особенности поведения корковых нейронов в разные стадии сна дают основание судить о нейрофизиологических механизмах его развития на нейронном уровне. Так, выключаемые из деятельности при засыпании нейроны, которые характеризуются полимодальностью, длинным латентным периодом и вариабельностью реакции, а также высокой частотой импульсации и повышенной возбудимостью, очевидно, имеют наиболее близкое отношение к осуществлению высших функций мозга. Можно предположить, что с их выключением связана потеря способности к активному восприятию окружающего и утрата сознания. Ограничение других функций мозга, возможно, обусловлено снижением возбудимости нейронов, замедляющих свои разряды, уменьшением информативности передаваемых сигналов, огрублением рабочих мозаик нейронной активности и перестройкой структуры импульсных потоков с преобладанием пачечного типа, характерного для восходящих таламокортикальных тормозных влияний.
Восстановление показателей нейронной активности в период ПС имело частичный характер. Можно предполагать, что это восстановление отражает растормаживание нейронных механизмов, имеющих отношение к явлениям, специфическим для ПС, как, например, быстрым движениям глаз или возникновениям сновидений. Однако для такого разграничения имеющихся сведений недостаточно, их могут дать дальнейшие исследования.
На рис. 45 приведена сводная схема расположения в структурах мозга систем, участвующих в регуляции уровня бодрствования и глубины сна. В соответствии с этой схемой последовательное включение нейрогуморальных механизмов, обусловливающих засыпание, смены стадий сна и пробуждение, можно представить по сигналам наступления темноты, прекращения деятельности и обстановки приготовления ко сну.
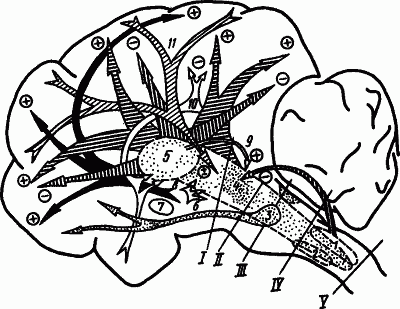 |
Рис. 45. Структуры мозга, участвующие в регуляции уровня бодрствования и глубины сна (по А.Н. Шеповальникову):
Условное и безусловное торможение, которое развивается на фоне изменения активности гормонов и метаболизма, а также ограничение сенсорной информации и умеренное утомление обусловливают угнетение внешней деятельности путем ослабления влияния активирующих десинхронизирующих систем (рис. 45,
Снижение тонуса активирующих систем помогает выявлению развивающейся к этому времени деятельности гипногенных синхронизирующих систем (рис. 45,
Нарастающая на протяжении десятков минут медленноволнового сна деятельность гипногенных систем достигает степени, при которой (пока неясно, каким образом) происходит быстрый переход к активами центров дорсомедиальных отделов моста (рис. 45,
Активность нейрогуморальных механизмов парадоксального сна постепенно уменьшается и может на протяжении нескольких минут снизиться настолько, что усилившиеся за это время влияния гипногенных систем вызовут быструю обратную перестройку деятельности мозга в состояние медленноволнового сна. Однако это состояние, хотя и более продолжительное, чем состояние парадоксального сна, через некоторое время снова сменится им. Такое циклическое чередование стадий сна длится всю ночь, причем к утру продолжительность стадий парадоксального сна обычно возрастает.
С утренним рассветом связаны такие изменения активности гормонов и обмена веществ, которые в суточной периодике означают переход к активной внешней деятельности в светлое время. Под влиянием этих изменений, а также нарастающих потоков сенсорной информации и завершения восстановительных процессов в накануне работавших до утомления структурах мозга происходит снижение тонуса гипногенных систем, что благоприятствует резкому возрастанию деятельности активирующих систем, особенно десинхронизирующего действия ретикулярной формации среднего мозга. Наступает пробуждение.
В разные стадии сна по-разному протекают процессы высшей нервной деятельности. Так, выработанная на словесную инструкцию двигательная реакция сжатия руки в кулак сопровождалась изменениями ЭЭГ и ЭМГ, наиболее выраженными в ПС. Описаны нарушения памяти при устранении ПС. Однако приводятся доказательства того, что этим не исключается консолидация долгосрочной памяти при выработке реакций пассивного избегания.
В управлении сном принимает участие наряду с нервной гуморальная регуляция. Из аминокислотных остатков выделен дельта-пептид сна, вызывающий дельта-волны и характерный для МС (М. Монье, Г. Шененбергер, 1975). Из мозга животных, лишенных сна, выделен фактор S, увеличивающий продолжительность МС. В опытах на животных и в наблюдениях на людях показана роль серотонина и норадреналина в процессах сна. На основе этих данных предложен следующий механизм развития и прекращения сна: подавление симпатической активности приводит к поверхностному сну, расходование серотонина соответствует глубокому МС, расходование норадреналина — ПС, возбуждение симпатической активности вызывает пробуждение.
По мере накопления факторов и наблюдений за сном человека и животных возникали разные теоретические представления о его природе.
Теория гипнотоксинов. Всем известное освежающее действие сна наводило на мысль, что за это время организм освобождается от накопившихся при дневной деятельности токсических продуктов обмена веществ, которые и вызывают сонное торможение нервных клеток мозга. Доказательством такого представления о химической природе сна служили опыты, в которых сыворотка крови долго не спавшей собаки, введенная другой собаке, вызывала у нее сон (Р. Лежандр, А. Пьерон, 1910). Однако скоро выяснилось, что это неспецифическое действие белков любой сыворотки крови. Кроме того, теории гипнотоксинов противоречит тот факт, что сон можно превозмочь интересным занятием. Самое наглядное опровержение химической теории сна дало наблюдение за сросшимися близнецами: при общем кровообращении, т.е. в одной и той же химической среде одна голова может спать, в то время как другая — бодрствовать.
Вместе с тем недавно было показано участие гуморальных факторов в развитии сна. Из крови животного, заснувшего в результате раздражения гипногенных зон таламуса, был получен уже упоминавшийся выше пептид, введение которого вызывало сон. Такой пептид дельта-сна, извлеченный из экстракта ткани ствола мозга, оказался активнее, чем приготовленный из ткани коры.
Теория центров сна. Эта теория ведет начало от клинических наблюдений за больными энцефалитом, вызывающим летаргический сон. У этих больных оказывался воспаленным определенный участок ствола мозга, который стали рассматривать как центр сна (К. Экономо, 1930). Предположение, что сон вызывается возбуждением специальных центров мозгового ствола, получило поддержку в опытах с раздражением структуры промежуточного мозга, под действием которого кошка укладывалась в характерную позу сна и засыпала (В. Гесс, 1933). Однако дальнейшие исследования показали, что такой результат можно получить, раздражая в определенном режиме стимуляции различные структуры мозга, что противоречило представлению о нервном центре, который должен был иметь определенную локализацию. Кроме того, клинические наблюдения показали, что патология сна не связана с определенной локализацией поражения мозга. Вместе с тем вопрос о центрах сна представляет существенный интерес.
Теория условного торможения. При изучении условных рефлексов в школе И.П. Павлова было обнаружено, что выработка различных видов условного торможения может приводить ко сну. Это наблюдали при угашении условного рефлекса, выработке дифференцировок, запаздывания, условного тормоза. Аналогичные обстоятельства вызывают сонливость у человека. Отсюда был сделан вывод, что «внутреннее торможение условных рефлексов и сон — один и тот же процесс» (И.П. Павлов, 1922). Особенность сна состоит в том, что торможение разливается, охватывая основные механизмы высших отделов мозга. Такое представление о природе сна объясняло накопившиеся к тому времени факты, в том числе противоречащие теориям гипнотоксинов и центров сна.
Теория деафферентации сенсорных систем. Основанием для этой теории явились факты развития глубокого сна у животных с выключенными основными путями поступления информации в большие полушария (путем перерезки мозгового ствола на уровне, предшествующем среднему мозгу). В подкрепление этой теории приводят описание больного, сохранившего из всех органов чувств только один глаз и одно ухо (этот больной засыпал, как только их закрывали), и эксперименты с хирургическим выключением у собаки зрения, слуха и обоняния, в результате чего она почти все время спала. С позиций теории условного торможения эти случаи считают проявлением «пассивного сна» (И.П. Павлов, 1932).
Теории неспецифических регуляторов сна и бодрствования. Особую роль в неспецифической регуляции функционального состояния высших отделов мозга играет восходящая активирующая система ретикулярной формации среднего мозга (Д. Моруцци, Г. Мэгун, 1949). Ее раздражение вызывает «реакцию пробуждения», десинхронизирует электроэнцефалограмму, повышает возбудимость коры мозга. Снижение влияний ретикулярной формации на кору приводит к возникновению «сонных потенциалов» и развитию сна. Этим объясняется глубокий непробудный сон после перерезок мозгового ствола перед средним мозгом. Если перерезка была сделана позади среднего мозга, такой сон не наступает. Возможно что патологический сон при летаргическом энцефалите был также результатом поражения ретикулярных структур среднего мозга. Силу активирующего действия ретикулярной формации демонстрирует опыт, в котором ее электрическая стимуляция пробуждает котенка, находящегося после длительного лишения сна в состоянии полной прострации и отсутствия реакций даже на болевые раздражения.
Действие активирующей, десинхронизирующей системы среднего мозга чередуется с действием тормозящих, синхронизирующих систем главным образом промежуточного мозга, отвечающих на низкочастотную стимуляцию. Именно здесь локализуются структуры, раздражение которых вызывает сон. Суточная периодичность природных факторов среды и состояния организма создают условия преобладания той или другой из этих систем, определяющих смену бодрствования и сна.
Современные представления о природе сна. Еще многое не ясно, однако современные успехи изучения сна все больше раскрывают сложный нейрогуморальный механизм его возникновения и развития. Основным в картине сна является потеря связи с внешним миром как результат затормаживания процессов деятельности высших отделов мозга. Такое торможение возникает и развивается в связи со многими обстоятельствами, среди которых важную роль играет суточная смена благоприятных и неблагоприятных условий существования (ночь для дневных и день для ночных животных), что обусловливает соответствующую периодичность всех функций организма. Поэтому на определяемое время суток и обстановку отхода ко сну в высших отделах мозга возникает условное торможение. Оно суммируется с безусловным торможением защитной неподвижности как компонентом инстинктивного поведения. К нему добавляется охранительное торможение, которое зависит от степени утомления корковых клеток во время дневной деятельности. Распространению тормозного процесса способствует ограничение афферентации при подготовке ко сну.
В то же время эндокринно-метаболические сдвиги, характерные для данного времени суток, изменяют соответствующим образом функциональное состояние нервных механизмов мозга. Такое действие оказывают главным образом гормоны гипофиза, щитовидной железы, надпочечников, а также медиаторы и другие биологически активные вещества. При этом холинэргические соединения способствуют активированию гипногенных синхронизирующих систем, в первую очередь медиальных таламических, бульбарных и структур основания мозга, возбуждение которых реципрокно и обратными связями через кору тормозит ретикулярную среднемозговую систему, поддерживающую состояние бодрствования. Развитие медленноволновой стадии (МС) сна связано с действием серотонина, накапливающегося главным образом в клетках средней линии мозгового ствола, а переход в стадию парадоксального (ПС) сна —норадреналина, аккумулирующегося в клетках покрышки моста. Получены сведения об участии в организации процессов сна специфических пептидов, моноаминов, гистамина, гамма-аминомасляной кислоты и других биологически активных веществ. Таким образом, физиологические механизмы сна включают в себя как нервные, так и гуморальные механизмы в их взаимодействии.
По поводу
Восстановление специфического метаболизма нервных клеток мозга, которое обеспечит его полноценную деятельность в состоянии бодрствования. И.П. Павлов (1935) полагал, что наступающее в течение напряженной дневной работы «истощение» корковых клеток вызывает сонное торможение, в течение которого восстанавливается их работоспособность. Когда оказалось, что в среднем нейроны коры продолжают разряжаться импульсами, было высказано предположение, что в «отдыхе» нуждается лишь небольшая их часть, так называемые обучающиеся нейроны, (
Приспособление к неблагоприятным условиям деятельности. Животные, ведущие дневной образ жизни, становятся беспомощными ночью, так как не ориентируются в темноте и могут стать легкой добычей ночных хищников. В свою очередь последние оказываются в аналогичном положении днем. Сон обеспечивает не только отдых, но и безопасность путем защитной неподвижности в укромном месте. Это один из видов инстинктивного приспособительного поведения.
Изучение нейрофизиологического механизма сна как сложнейшего безусловного рефлекса выявило свойства его возникновения и протекания, а также участие в этом определенных структур головного мозга (рис. 46). Дальнейшие исследования выявили усложнение и своеобразие форм сна в эволюции, отражающие особенности уровня развития нервной системы и экологию вида.
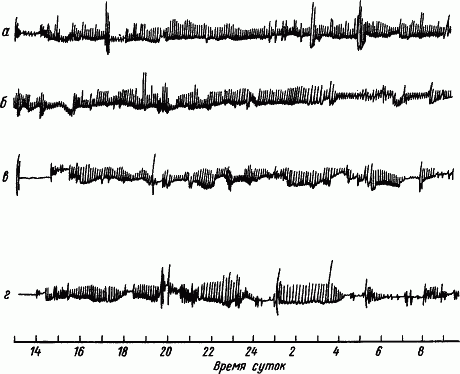 |
Рис. 46. Фрагменты непрерывной регистрации движений бесполушарного голубя, находящегося при постоянном освещении 100 суток в состоянии бодрствования (по Н.А. Рожанскому):
Значение сна как акта инстинктивной деятельности рассматривали с точки зрения двух ее компонентов: формирования потребности и ее удовлетворения. Одни исследователи считают, что оба компонента реализуются в ПС сна (Т.Н. Ониани, 1976), другие — связывают потребность с МС, а ее удовлетворение с ПС (Г. Фогель, 1979).
Упорядочение процессов обработки и хранения информации. Значение сна для состояния памяти понимают двояко. Ряд ученых считают, что происходит устранение накопившейся за день «ненужной» информации, наступает «дезынтеграция памяти», такую подготовку мозга к восприятиям следующего дня сравнивают со стиранием информации в ячейках памяти запоминающего устройства вычислительной машины. Другие, наоборот, считают, что во время сна происходит консолидация памяти, переход кратковременной в долговременную. Высказывают также предположение о переработке во время сна информации, которую мозг не успел переработать в течение дня. В последние годы сон стали рассматривать как механизм «психологической защиты» от внутренних, обычно неосознаваемых конфликтных ситуаций, например неосуществимых желаний и пр. Действие этого механизма связывают с ПС сна и возникающими в это время сновидениями. Такой подход к значению сна близок развиваемой в свое время концепции о скрытых в подсознании влечениях, которые проявляются в сновидениях и могут быть обнаружены психоанализом (З. Фрейд, 1933). Сами сновидения оценивают как средство снятия психического напряжения путем имитации решения «текущих эмоциональных проблем». Современные физиологические и клинические исследования указывают на большое функциональное значение сновидений. Для расшифровки сновидений разрабатывают своеобразные «грамматики сна».
Восстановление согласованности временного течения функций организма. Бесчисленное множество биохимических реакций выстраивается в сложную систему обеспечения функций клеток, тканей и органов. Согласование во времени этих взаимосвязанных периодически изменяющихся функций является необходимым условием нормальной жизни организма. Нарушения согласования ведут к тяжелым явлениям десинхроноза. Такое рассогласование со всеми его последствиями возникает, например, при сдвинутых режимах сна, перелетах с пересечением часовых поясов, когда внутренние ритмы течения функций приходят в противоречие с внешними периодами смены дня и ночи. Однако и в обычных условиях активной деятельности днем многочисленные воздействия и ответные реакции в какой-то степени нарушают плавное течение ритмики тех или иных функций. Предполагают, что становление их согласования происходит во время сна, когда внешние воздействия выключаются. Именно поэтому одним из симптомов десинхроноза является повышенная потребность в сне.
Длительное торможение процессов высшей нервной деятельности, сопровождаемое резким снижением уровня вегетативных функций, наступает у некоторых видов животных как приспособление к существованию в неблагоприятные сезоны года. Оно получило название
Многие животные, обитающие в местности, где зимой они лишаются источников питания и не могут вести активный образ жизни, впадают на это время в
Последовательность затормаживания мозговых структур видна на рис. 47. Обращает на себя внимание тот факт, что уже после прекращения активности в неокортексе в гиппокампе наблюдается «взрыв» деятельности. По поводу влияния спячки на условные рефлексы существуют противоречивые данные. Так, описана потеря во время спячки выработанных ранее условных рефлексов у хомяков и ежей, но сохранение их у сусликов. Однако некоторые безусловные рефлексы во время спячки оказываются усиленными. К ним относятся сложные рефлексы поддержания специфической позы спящего животного, в том числе проприоцептивные и лабиринтные (так, если летучую мышь, которая во время спячки висит вниз головой, перевернуть, то она переползает так, чтобы снова очутиться в прежней позе).
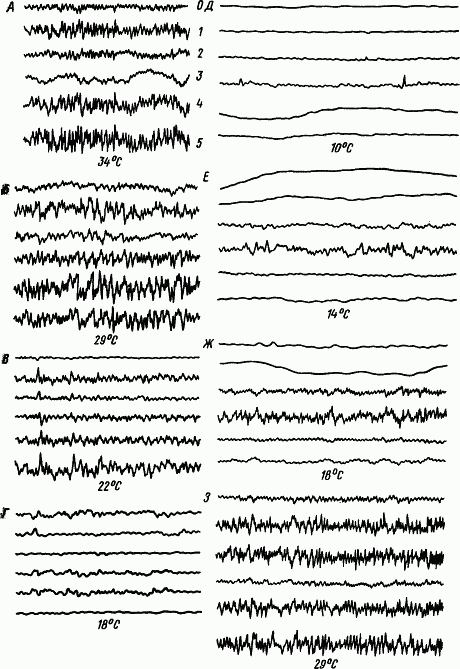 |
Рис. 47. Электрическая активность разных отделов головного мозга европейского ежа при бодрствовании
цифрами показаны отведения,
Наиболее яркое и удивительное изменение функций организма при такой спячке состоит в значительном сокращении обмена веществ. Исследование скорости включения меченого фосфора (Р98) в ткани мозга показало его резчайшее падение во время зимней спячки суслика. Примечательно, что наиболее угнетенным оказывается синтез РНК. Траты энергии уменьшаются во много раз по сравнению с бодрствованием. Температура тела снижается почти до уровня, установившегося в норе или берлоге.
Описано крайнее замедление частоты дыхания при наступлении зимней спячки по сравнению с периодом бодрствования: с 40–50 вдохов/мин до 1–6 у ежей, с 25 до 4–5 у сурков, а у орешниковой сони наблюдались даже 5–10-минутные остановки дыхания. Падают величины основных показателей кровообращения. Так, частота сердечных сокращений снижается у ежа с 230 до 24 ударов/мин, объем крови, выбрасываемой в 1 мин сердцем суслика, уменьшается в 60 раз, время кругооборота крови возрастает в десятки раз, артериальное кровяное давление становится в 5–6 раз ниже. Глубокие перестройки происходят в эндокринной системе и процессах тканевого метаболизма, изменяется течение всех вегетативных функций. Следует заметить, что регуляция этих функций у большинства зимоспящих животных уже в состоянии бодрствования обнаруживает особенности, создающие благоприятные условия для развития явлений спячки. Так, и в состоянии бодрствования температура тела у них оказывается ниже, чем у животных, не впадающих в зимнюю спячку.
Многие обитатели пустынь и полупустынь в засушливое время года при высокой температуре среды, высыхании водоемов и истощении растительности погружаются в
Нервный механизм возникновения и развития сезонных спячек включает в себя как условные, так и безусловные рефлексы, перестройки ряда процессов, прежде всего энергетического обмена и терморегуляции. Эти перестройки происходят под влиянием климатических и биоценотических сигналов на фоне их сезонной периодики, сложившейся в эволюции. Исполнительными механизмами являются терморегуляторные центры мозгового отдела и эндокринные регуляторы обмена веществ. О значении факторов питания свидетельствуют эксперименты, в которых полное водное голодание вызвало у малого суслика летнюю спячку.
Своеобразным сноподобным состоянием является так называемый
Сходные явления каталептической неподвижности наблюдали в лабораториях И.П. Павлова, как переходное состояние между бодрствованием и сном, вызванным иррадиацией внутреннего торможения в коре при исследовании условных рефлексов у собак. Когда в результате однообразия экспериментальной обстановки и действия отрицательных раздражителей торможение начинало распространяться по структурам мозга, то вначале выключались произвольные движения (реакция двигательной коры). При этом действовали стволовые центры сохранения данной позы слюноотделительных рефлексов. Затем наступала каталептическая неподвижность: собака застывала в станке, не брала пищу, но выделяла на ее вид слюну. Дальнейшее распространение торможения на структуры мозгового ствола вызывало ряд промежуточных состояний постепенной потери позы, расслабления мускулатуры, наконец, прекращения выделения слюны на пищу, наступал полный сон. Такие промежуточные состояния можно было видеть в динамике показателей условных слюноотделительных рефлексов и по аналогии со стадиями
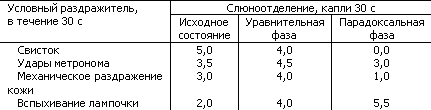
В табл. 12 показаны результаты одного из опытов, когда в исходном бодром состоянии животного величина рефлекса зависит от силы условного раздражителя в ряду: свисток, удары метронома, механическое раздражение кожи, вспыхивание лампочки. Однако после развития напряженного дифференцировочного торможения, «столкнувшегося» с возбуждением, возникли переходные состояния в виде гипнотических уравнительной и парадоксальной фаз.
Таблица 12. Гипнотические фазы переходных состояний условно-рефлекторной деятельности (по И.П. Разенкову)
 |
С давних пор вызывали и вызывают у всех большой интерес явления
Внушения при гипнозе приобретают особую силу потому, что они не контролируются всеми остальными заторможенными механизмами сознания. Загипнотизированному можно внушить, что он обедает, и он будет с аппетитом черпать ложкой из пустой тарелки. При этом у него «слюнки текут», начинает выделяться желудочный сок и изменяется состав крови. Гипнозу очень легко поддаются люди с сильной уравновешенной нервной системой, способные к самоорганизации и аутотренингу, а также с ослабленной нервной системой, в которой возбуждение скорее переходит в торможение. В то же время гипнотический сон является для таких людей лечебным средством, снимающим нагрузку, и может заменять снотворные при терапии сном. Он применяется также при лечении некоторых нервных заболеваний, алкоголизма и для мобилизации резервных сил организма больного путем внушения ему веры в выздоровление.
С психологической точки зрения гипноз был образно определен как
| © 2025 Библиотека RealLib.org (support [a t] reallib.org) |