"Чувства животных" - читать интересную книгу автора (Бертон Роберт)
ГЛАВА 2 Тревожно прислушиваясь…
Прежде чем описывать механизмы функционирования уха, необходимо дать ясное представление о том, что такое звук.
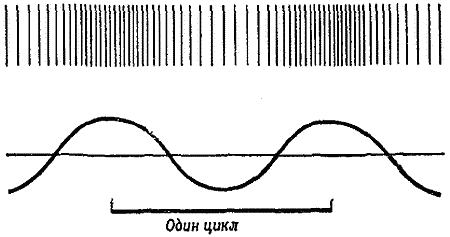
Стоит лишь ударить рукой по струнам гитары или по камертону, как они начинают колебаться. Вибрирующая струна передает колебания частицам окружающего воздуха, а они в свою очередь — соседним частицам, создавая таким образом серию чередующихся сжатий и разрежений с усилением и ослаблением давления воздуха. Эти движения частиц графически изображаются в виде последовательности волн, вершины которых соответствуют сжатиям, а впадины между ними — разрежениям. Скорость движения этих волн в данной среде и есть скорость звука. В воздухе на уровне моря звук распространяется со скоростью около 1200 км/ч, а о самолетах, которые летают быстрее, говорят, что они преодолевают звуковой барьер. На больших высотах, где воздух разрежен, скорость звука падает, и самолеты преодолевают звуковой барьер на значительно меньшей скорости, чем 1200 км/ч. Напротив, в более плотной среде, например в воде, звук распространяется — быстрее, а в скале — еще быстрее.
Число волн, проходящих в секунду через какую-либо точку пространства, называется
 |
Ухо человека или какого-либо животного воспринимает звук лишь в ограниченном диапазоне частот или длин волн. Волны давления с частотой ниже 20 Гц не воспринимаются нашим ухом как звуки, а ощущаются как вибрации. Вместе с тем колебания с частотой выше 20 000 Гц (так называемые
Верхняя граница слуха у различных людей не одинакова, и обычно дети могут слышать более высокие звуки, чем взрослые. В качестве примера приведем рассказ о маленьком четырехлетнем мальчике, который ночью разбудил своих родителей, устроив большой переполох. Поскольку он редко просыпался по ночам, родители решили выяснить, в чем дело. Судя по рассказу мальчика, по комнате что-то летало, издавая писк. Ничего не обнаружив, родители попытались успокоить ребенка, но он еще больше разволновался и упорно настаивал на том, что в комнате что-то есть. Вдруг ребенок закричал, что
Следует отметить, что в данном случае летучая мышь посылала отнюдь не ультразвуковые импульсы, используемые для эхолокации (гл. 4). Это был обычный писк, которым пользуются мыши для общения друг с другом. Приведенный рассказ ясно показывает, что мы не можем допустить даже мысли, будто один человек воспринимает те же звуки, запахи или зрительные сигналы, что и другой. И когда мы изучаем животных, необходимо четко представлять, что диапазон их восприятия мира совершенно отличен от нашего.
Другой характеристикой звуковых волн является
Звуковые волны, порожденные вибрирующей струной гитары, камертоном или каким-либо другим источником, собираются наружным ухом, или ушной раковиной (фиг. 5,7), и направляются по слуховому проходу к барабанной перепонке. Она представляет собой мембрану диаметром 1 см, которая колеблется, когда о нее ударяются звуковые волны, и служит, таким образом, первым звеном преобразователя. Эти колебания усиливаются и передаются к рецепторным клеткам внутреннего уха с помощью среднего уха.
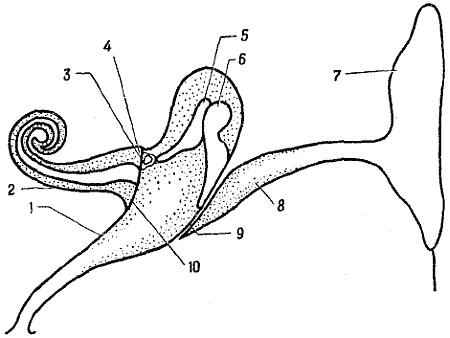 |
Среднее ухо представляет собой камеру, называемую
Чтобы предохранить внутреннее ухо от повреждения при усилении слишком громкого звука, к молоточку и наковальне прикреплены особые мышцы, которые, сокращаясь, отводят эти косточки от барабанной перепонки и овального окна. При этом звук может проходить во внутреннее ухо, но его интенсивность значительно ослабляется.
Внутреннее ухо состоит из двух заполненных жидкостью образований, находящихся внутри височной кости.
Уже ранние эксперименты показали, что базилярная мембрана играет очень важную роль в восприятии волн давления и преобразовании их в нервные импульсы. Непосредственно над ней лежит
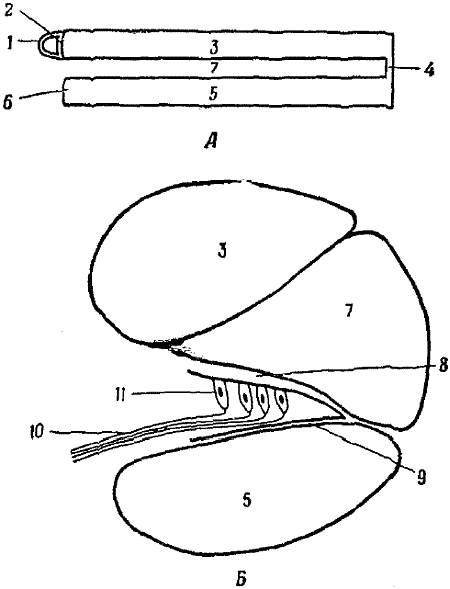 |
Однако эту теорию легко опровергли опыты с перерезкой волокон базилярной мембраны. Перерезанные очень острым скальпелем концы не отскакивали в стороны, а значит, волокна не были напряжены и, следовательно, не могли резонировать. Более близкая к истине, но и более сложная теория была разработана совсем недавно. В экспериментах использовали маленький поршень, помещенный на место стремечка. Движениями этого поршня можно было механически управлять таким образом, чтобы улитке посылались волны известной частоты. Амплитуда и скорость волн увеличивались по мере распространения их по улитке, пока не достигали максимума, и затем волны замирали. Амплитуда и скорость колебаний различной частоты, создаваемых поршнем, достигали максимума в различных местах базилярной мембраны. Высокочастотные колебания достигали максимума вблизи поршня, а низкочастотные — около вершины улитки, на другом конце мембраны. Это подсказало возможный прямой способ механического анализа частот при участии базилярной мембраны. Но анализ этот не прост, так как даже чистый тон одной частоты заставляет вибрировать значительный участок мембраны.
Во время этих экспериментов было обнаружено, что базилярная мембрана несет небольшой электрический заряд, сходный с зарядом, который несет нервное окончание в тельце Пачини. В отсутствие вибраций величина заряда постоянна, но, когда проходят волны давления, она начинает колебаться. Если надавить на базилярную мембрану, заряд увеличивается, а если мембрану приподнять, то он ослабевает. Такие изменения постоянного заряда возникают в результате изгибания волосковых клеток, когда колеблющаяся базилярная мембрана также заставляет их колебаться. Эти изменения заряда точно соответствуют как по частоте, так и по амплитуде изменениям давления в жидкости улитки, которые создаются вибрирующим поршнем. Здесь происходит то же самое, что и в микрофоне и в пачиниевом тельце (гл. 1). Описанные выше меняющиеся заряды называются
Ухо чувствительно не только к частоте и громкости звуков; оно может также определять, откуда приходят звуки. Такие животные, как олень или осел, имеющие подвижные ушные раковины, устанавливают местоположение источника звука, поворачивая их до тех пор, пока звук не станет возможно более громким. Этот же принцип используется при навигации по радиомаякам: приемная антенна поворачивается до тех пор, пока сигнал маяка не становится максимальным; в этом случае направление оси антенны совпадает с направлением, откуда приходят сигналы.
Стоит только понаблюдать за козой (фото 1) [1] кошкой или собакой, чтобы увидеть, как они пользуются своими ушными раковинами для улавливания звука. Если мимо поля, где пасется коза, проезжает машина, уши козы поворачиваются по кругу, прослеживая за движением машины по дороге. А когда приближается второй автомобиль, коза поворачивает одно ухо назад, чтобы сосредоточить внимание на обоих звуках сразу, и если двигатель одной из машин вдруг «выстрелит», коза запрядет ушами, защищаясь от шума.
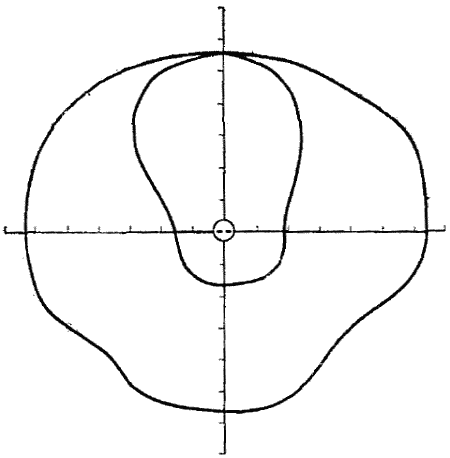 |
Простой опыт с моделью уха козы показывает, как ушная раковина увеличивает способность животного более точно определять направление, в котором находится источник звука. Такая модель с вмонтированным в нее микрофоном устанавливается на вращающемся столике на некотором расстоянии от источника звука. При вращении столика регистрируется сила тока, поступающего от микрофона. Результаты этих опытов представлены в виде диаграммы на фиг. 7. Ухо козы наиболее чувствительно к звукам, источник которых находится несколько сбоку от животного, и плохо воспринимает звуки, приходящие сзади.
В течение долгого времени ушные раковины человека наряду с аппендиксом приводили в качестве примера органов, потерявших свои первоначальные функции и сохранившихся лишь в качестве рудиментов. Считалось, что когда-то они были похожи на уши козы или оленя. Некоторые из нас все еще могут двигать ушами, но, конечно, не настолько хорошо, чтобы защищаться таким образом от неприятных звуков. Несколько лет назад было обнаружено, что ушные раковины человека не так бесполезны, как это казалось. Они, вероятно, не столь эффективны в качестве собирающих воронок, помогающих слышать слабые звуки, но играют важную роль в определении направления, откуда приходят звуки. В 1968 году были опубликованы результаты исследований, проведенных на людях; эти исследования служат хорошим примером того, как совсем простые тесты или наблюдения могут показать, что наши прежние представления, основанные часто лишь на предположениях, нередко оказываются совершенно неверными. Первыми шагами, которые направили нас по верному пути, были исследования, показавшие, что при деформации ушной раковины человеку становится трудно точно определять положение источника звука. Если бы ушная раковина служила просто коллектором звуков, наподобие слуховой трубки, то ее деформация привела бы только к ослаблению способности уха слышать очень тихие звуки; оказалось, однако, что ушная раковина предназначена еще и для каких-то других целей.
Первоначально возможности таких экспериментов были ограничены, поскольку ушные раковины человека можно деформировать лишь до известного предела, не рискуя вызвать необратимые повреждения. Кроме того, ушные раковины человека невозможно заменить ушными раковинами других конфигураций и оценить их эффект. Проблема была решена, когда изготовили модели ушных раковин человека с вмонтированными в слуховой проход микрофонами. Воспринятые такими моделями звуки передавались затем экспериментатору с помощью специальных наушников. Опыты показали, что хрящевые бугорки в наших ушных раковинах действуют подобно дросселям, задерживая звук, когда он входит в ухо. Величина этой задержки зависит от угла, характеризующего направление на источник звука. Оценивая разницу во времени прихода звука к каждому уху, мозг способен определить местонахождение источника звука. Если этот источник расположен прямо впереди или сзади, звук приходит в оба уха одновременно; если он находится слева, то требуется больше времени, чтобы звук достиг правого уха, и т. д.
Существуют два других способа, с помощью которых животные определяют местонахождение источника звука; в зависимости от размеров головы и длины звуковой волны различные животные пользуются этими способами в большей или меньшей степени. Даже в отсутствие ушной раковины ухо обладает разной чувствительностью к звукам, приходящим с разных сторон (фиг. 7). Используя оба уха, можно определить местонахождение источника звука с помощью стереоскопического слуха, точно так же как определяется расстояние с помощью стереоскопического зрения. Положение источника звука оценивается на основе различной громкости звука в каждом ухе или времени его прихода в каждое ухо. Если частота звука превышает 15 кГц, голова служит для него своего рода препятствием, подобно волнорезу, на задней стороне которого звуковые волны отсутствуют. Вследствие этого ухо, расположенное ближе к источнику звука, регистрирует значительно более громкие сигналы, чем то, которое находится дальше от него. Кроме того, оценивается и разница во времени прихода звуковой волны к каждому уху. Если источник звука расположен на одинаковом расстоянии от каждого уха, то звуковая волна достигает обоих ушей одновременно. Если голова повернута всего лишь на 5° в сторону, разница во времени составляет 0,00004 с, а при повороте на 90° эта разница составит 0,0005 с. При наличии определенной разницы в громкости звука и во времени его прихода к ушам мозг способен уловить небольшие различия между сигналами, приходящими от каждого уха. Он может различать сигналы, отстоящие друг от друга во времени не менее чем на 0,0001 с, что представляет собой сравнительно большой временной интервал. Этот недостаток в какой-то степени корректирует дроссельная система, имеющаяся в наших ушах, или движения головы из стороны в сторону, в результате чего каждое ухо по изменению громкости звука может очень точно определять местонахождение его источника; точно так же поступает коза, когда двигает ушными раковинами. Но даже все эти приспособления не очень эффективны при определении местонахождения источника звука, и обнаружить его обычно помогает зрение.
Иначе обстоит дело у сипухи. До недавнего времени считалось, что совы охотятся исключительно с помощью зрения. У них большие глаза и, по-видимому, достаточно хорошее зрение, чтобы они могли охотиться даже тогда, когда луна и звезды скрыты облаками. Проведенные в последние годы опыты показали, что на самом деле сипуха способна обнаруживать свою жертву с помощью одного только слуха, набрасываясь на движущуюся мишень с поразительной точностью. Опыты проводили в темной светонепроницаемой комнате. В первой серии экспериментов пол в комнате покрывали сухими листьями и выпускали туда мышь. Можно было слышать, как мышь пробирается сквозь листья и затем останавливается. Тотчас же было слышно, как сипуха покидает свой насест, и глухой звук возвещал о том, что она уже на полу. Когда в комнате включали свет, видели, что мышь в когтях у совы. Дальнейшие опыты показали, что сова попадала в цель гораздо чаще, чем промахивалась, а если и промахивалась, то мыши удавалось ускользнуть только чудом.
Однако без тщательной проверки эти опыты еще нельзя было считать доказательством того, что сова определяет местоположение жертвы по создаваемому мышью шуму. Хотя светонепроницаемость комнаты и лишала сову возможности пользоваться зрением, она могла обнаруживать мышь с помощью эхолокации, по запаху или теплу, излучаемому ее телом, так же как змеи обнаруживают тепло, излучаемое телами животных (гл. 10). Эти возможности были исключены в серии контрольных опытов, в которых по полу на веревочке протаскивали бумажный шарик. Сова так же уверенно бросалась и схватывала бумагу; следовательно, для обнаружения своей жертвы она пользовалась только слухом. Чтобы доказать это окончательно, мышь выпускали на голый пол, на котором она не производила никакого шума, и в этом случае сова оказывалась совершенно неспособной обнаружить ее.
Тем не менее эти эксперименты еще не доказывают однозначно, что сипухи охотятся только с помощью слуха. Их зрение в десять, а то и в сто раз лучше нашего, и вполне вероятно, что сипухи пользуются им, когда охотятся в сумерках или при свете луны. Если бы сипухи полагались только на зрение, то в облачные или безлунные ночи они оказались бы на голодном пайке; поэтому хороший слух служит им ценным запасным средством для обнаружения жертвы. Нельзя считать, что у всех сов такой же хороший слух или такое же хорошее зрение, как у сипухи. Домовый сыч и американская кроличья сова охотятся днем. Вряд ли они очень часто пользуются слухом, хотя в темноте они видят не лучше, чем мы с вами.
Способность сипухи и, как теперь известно, других охотящихся ночью сов так точно определять местоположение своей жертвы по слуху отчасти обусловлена строением их уха. Ухо этих птиц построено по такому же принципу, что и наше, но улитка у них прямая, а не спиралевидная, и три косточки среднего уха заменены одной, называемой
У сов большая барабанная перепонка, так что прилегающая к ней колонка передает значительное количество звуковой энергии к овальному окну улитки, размеры которого относительно малы. Поэтому давление звука усиливается теоретически в 40 раз, а не в 18, как в нашем ухе. У большинства птиц колонка примыкает к центру барабанной перепонки, а у сов она расположена несколько эксцентрично, что обеспечивает дополнительное усиление давления. Барабанную перепонку можно рассматривать как рычаг. «Точкой» опоры при этом служит периметр перепонки, где ее края прикрепляются к кости. Когда барабанная перепонка колеблется, ее центр выпячивается больше, чем края; при этом она действует как рычаг, передвигая колонку вперед и назад. Сместившись от центра и оказавшись ближе к точке опоры, колонка передвигается хоть и не так далеко, как если бы она находилась в центре, но зато с большей силой. Таким образом происходит усиление попадающего в ухо совы звука, прежде чем он будет передан внутреннему уху.
Однако сове мало иметь острый слух — она должна еще знать, откуда доносится слабый шорох, производимый мышью. Другими словами, она должна уметь определять местонахождение источника звука. Сова пользуется тем же средством, что и мы с вами, различая громкость приходящих в каждое ухо звуков и оценивая разницу во времени их прихода.
Мелкие птицы, например воробьи, садовые славки и др., не могут пользоваться вторым способом, поскольку у них очень маленькие головы и интервал между приходом звуковой волны в одно и другое ухо слишком мал, чтобы его можно было уловить. У сов головы значительно больше, благодаря чему и интервал достаточно велик, чтобы сова его ощутила.
Для определения местонахождения источника звука могут также использоваться и различия в громкости звука, слышимого каждым ухом; эксперименты, в которых применялся вставленный в ухо совы микрофон, показали, что оно, подобно уху козы, более чувствительно к звукам, идущим спереди, чем к звукам, приходящим откуда-либо еще. Это происходит потому, что у сипухи вокруг каждого уха имеется кожная складка, наподобие ушной раковины, которая концентрирует звук. Более того, хотя уши расположены симметрично по бокам головы, эти складки занимают несимметричное положение, благодаря чему увеличивается разница в громкости звуков, достигающих обоих ушей, и удается более точно определить локализацию их источника. Поворачивая голову, сова определяет, при каком положении головы громкость звука, приходящего в каждое ухо, максимальна. Таким образом, она фактически определяет два пеленга на источник звука.
Что же может противопоставить мышь способности совы находить и почти без промаха схватывать свою жертву? Убивает ли сова без разбора любую мышь, так что каждая мышь имеет одинаковые шансы быть съеденной, или же существует известная избирательность, как можно было бы ожидать, если верна теория о выживании наиболее приспособленных? Исследования взаимоотношений хищника и жертвы показывают, что хищники, как правило, не убивают случайных особей. Большая часть их попыток поймать жертву завершается неудачей, а в случае успешной охоты жертвой обычно становятся животные молодые, слабые или оказавшиеся в неблагоприятных условиях, что увеличивает их шансы попасть в лапы хищника. Мыши, которых выпускали в комнате с сипухой, были в явно невыгодном положении. Они попадали в незнакомое место, которое им нужно было исследовать. Если бы мышам дали время освоиться, поймать их было бы труднее.
Опыт с сипухой показал, что сова может охотиться, руководствуясь только слухом; однако следует учитывать, что выводы, полученные в экспериментах, часто вводят нас в заблуждение, если мы без всяких оговорок применяем их к реальной ситуации. На воле мыши, полевки и другие мелкие животные не создают сильного шума. У каждой мыши есть своя система трасс, проложенных в траве или подлеске, по которым она неслышно бегает, находясь в относительной безопасности. Можно притаиться на поросшей травой лужайке, прекрасно зная, что там есть мыши, и не уловить ни малейших признаков их передвижений.
Чаще всего попадают в когти хищника те мыши, которые оказываются вне системы своих трасс. Это молодые мыши, еще не освоившие мест обитания, или те, которые вынуждены из-за перенаселенности покинуть привычную сеть дорог. У них есть единственный способ защиты — замереть при малейшем признаке опасности. Однако опыты показали, что к тому времени, когда мышь обнаружит опасность, сова, как правило, уже знает, где находится ее жертва. Кроме того, у совы есть другое преимущество, поскольку она летает почти бесшумно. Маховые перья ночных сов имеют мягкие опахала, благодаря чему значительно ослабляется шум крыльев, рассекающих воздух. Мыши остается надеяться только на то, что сова криком выдаст свое присутствие. Раньше считали, что сова кричит для того, чтобы напугать мышь и выгнать ее на открытое место, однако, как оказалось, крик совы вызывает скорее обратный эффект. Услышав этот крик, мышь замирает.
Не все мелкие животные живут в таких местах, где можно легко спрятаться от сов в густом подлеске. Некоторые обитают в пустынных или полупустынных местностях, где трудно найти укрытие. Замечательная особенность многих мелких пустынных животных, таких, как слоновая землеройка, африканский тушканчик и кенгуровая крыса, обитающая в сухих местностях на западе США и в Мексике, состоит в том, что у них очень большие барабанные полости, образованные куполообразными косточками, которые расположены позади нижней челюсти и окружают среднее ухо. Эта особенность характерна также и для более крупных обитателей пустыни, например для антилопы мендес, живущей в глубинных районах пустыни Сахара. Уже давно предполагали, что такие большие барабанные полости связаны с наличием острого слуха. Некоторые опыты, проведенные на кенгуровых крысах, очень четко показали, что и в самом деле эти животные обладают хорошим слухом, который предупреждает их о приближении врагов.
Кенгуровая крыса — и не кенгуру, и не крыса. Это небольшое животное, которое похоже на мышь и одновременно очень напоминает кенгуру. У нее большие и сильные задние лапы, с помощью которых она прыжками передвигается по земле, используя для поддержания равновесия свой длинный хвост. Кенгуровая крыса — ночное животное; в темноте она выходит на поиски пищи (зерна), которую собирает в защечные мешки и уносит про запас в свою нору.
Зоологи изучали кенгуровую крысу главным образом из-за ее способности питаться сухой пищей и обходиться без воды, но нас интересуют ее большие тонкие как бумага ушные кости, которые образуют полость среднего уха. Две эти полости по объему даже больше, чем мозг животного. Внутри барабанной полости обнаружены некоторые из тех приспособлений, обеспечивающих повышенную чувствительность, которые имеются и в ухе сипухи: у кенгуровой крысы большая барабанная перепонка, а стремечко плотно примыкает к очень маленькому овальному окну. Сравнение площадей барабанной перепонки и овального окна показывает, что среднее ухо усиливает звуки почти в сто раз.
Большой объем барабанной полости у кенгуровых крыс также повышает чувствительность их уха. Это было проверено на анестезированных животных, в улитку которых аккуратно вводили маленькие электроды. Затем на крыс воздействовали звуками различной частоты и с помощью электродов регистрировали кохлеарные микрофонные потенциалы. Было обнаружено, что кенгуровые крысы проявляют особую чувствительность к звуковым колебаниям частотой от 1000 до 3000 Гц. Когда барабанные полости чем-нибудь заполняли, чувствительность ушей сразу резко падала, хотя крысы продолжали реагировать на звуки того же диапазона частот.
Следовательно, большие барабанные полости должны улучшать слух кенгуровой крысы. Причина этого, по-видимому, заключается в том, что при нормальных размерах среднего уха колебания барабанной перепонки ослабляются из-за давления воздуха в расположенной позади нее барабанной полости. Если барабанная полость мала, то барабанная перепонка, вдавливаясь в полость, быстро создает в ней повышение давления, которое противодействует этому движению, а если барабанная полость велика, такого противодействия нет и барабанная перепонка может свободно колебаться. Это особенно справедливо в отношении звуков низкой частоты. Низкочастотные колебания вызывают наиболее сильные колебания барабанной перепонки, а именно к колебаниям низкой частоты более всего и чувствительны кенгуровые крысы,
Следующий этап этих экспериментов заключался в том, чтобы выяснить, имеет ли какое-нибудь значение для кенгуровой крысы чрезвычайно высокая чувствительность ее уха к звуковым колебаниям. Достаточно ли оно чувствительно, чтобы предупредить животное о появлении хищников? Кенгуровые крысы ведут ночной образ жизни; следовательно, их смертельными врагами являются ночные хищники. В пустынях США это совы и гремучие змеи. Поэтому были проведены эксперименты, подобные ранее описанным опытам с сипухой и мышью, но на этот раз кенгуровых крыс заставляли вступать в единоборство с сипухой и гремучей змеей. Оба хищника могут охотиться в темноте: сипуха, как было доказано, с помощью слуха, а гремучая змея — воспринимая тепло, излучаемое телом жертвы. Сначала кенгуровую крысу помещали в клетку с совой. Пользуясь красным светом, которого не видят крысы, можно было наблюдать сцену охоты в «полной темноте». Оказалось, что при каждом нападении совы кенгуровая крыса отпрыгивает в сторону. Это было захватывающее зрелище: когда сипуха вот-вот должна была схватить ничего, казалось бы, не подозревающую кенгуровую крысу, крыса быстро подпрыгивала и приземлялась в 25…30 см от того места, где она только что находилась, предоставляя сове хватать лишь землю. То же самое происходило и при нападении змеи. Наконец, чтобы доказать, как важны для кенгуровой крысы такие большие барабанные полости, проводили следующий опыт: эти полости заполняли пластилином, и кенгуровые крысы теперь до последнего мгновения так и оставались в полном неведении о грозящей им опасности.
Предстояло решить еще один последний вопрос. Перья сов имеют мягкие опахала, так что эти птицы летают бесшумно. Как же в таком случае кенгуровая крыса обнаруживает сову? Когда проводились описанные выше эксперименты, включали магнитофон, установив максимальное усиление. Тщательный анализ записи показал, что, когда сова нападает на свою жертву, слышится слабый шуршащий звук с частотой до 1200 Гц. Змея при нападении также производит звуки частотой до 2000 Гц. Эти частоты относятся к тому диапазону, в котором наиболее чувствительно ухо кенгуровой крысы, хотя их интенсивность настолько мала, что она соответствует пределу чувствительности магнитофона.
Итак, в процессе эволюции сипуха выработала способ обнаруживать свою жертву и скрывать свое присутствие. Ее жертвы научились прятаться или выработали способы быстрого обнаружения опасности. Однако ни в одном случае эти способы не являются совершенными: иногда мышь слишком медлительна, а иногда сове приходится поголодать, так что между хищником и жертвой существует некое равновесие.
| © 2024 Библиотека RealLib.org (support [a t] reallib.org) |