"Карл Х.Прибрам. Языки мозга (Экспериментальные парадоксы и принципы нейропсихологии) (fb2) " - читать интересную книгу автора (К.ПРИБРАМ)
Глава IV
Как можно охарактеризовать формы тех временных нейро-электрических конфигураций (и отношений между ними), которые делают возможным существование самих механизмов кратковременной памяти, механизмов, на какие мы опираемся, когда имеем дело, например, с конструкцией немецкого предложения с глаголом на конце? Изучение условий, влияющих на человеческую память, продемонстрировало несколько пренебрежительное отношение к проблеме структур; если что-либо запоминается,, то это объяснялось в значительной мере формой материала и контекстом, в котором он предъявлялся. Мы же обращаемся к проблеме замещения одной конфигурации другой, которое возникает в результате определенных операций в нервной системе. На техническом языке – это проблема преобразований или функций передачи, что делает возможным кодирование и перекодирование.
Операции кодирования постоянно имеют место в нервной системе. Физическая энергия воспринимается рецепторами и трансформируется в нервные импульсы. Эти импульсы в свою очередь достигают синаптических сетей, где дискретные сигналы кодируются в микроструктуры медленных потенциалов. Для того чтобы кодирование было эффективным и могло быть использовано мозгом, необходимо наличие декодирующей операции, благодаря которой было бы возможно восстановление нервных импульсов и сохранение, таким образом, предварительно закодированной информации.
Для описания операций кодирования, происходящих в нервной системе, можно использовать два класса функций передачи.
Одни преобразования допускают достаточно простой расчет соответствия между кодами. Такой расчет делает возможным декодирование закодированной формы, и наоборот. Благодаря однозначному соответствию между цифрами кода между ними устанавливается нечто вроде обратимости, вторичный функциональный изоморфизм.
Второй класс функций передачи целиком неизоморфный и условный. Преобразования в этом классе необратимы, пока нет ключа для дешифровки такого кода.
Обратимые трансформации возникают в физико-оптических системах, в звуковых преобразователях и, как мы покажем, в закодированных нейронных конфигурациях микроструктуры медленных потенциалов мозга. Необратимые преобразования имеют место в обычных языках, состоящих из условных алфавитных комбинаций, в азбуке Морзе, в телефонной и телевизионной передаче сигналов и т. п. В нервной системе, как будет подробно рассмотрено в последующих главах, имеют место необратимые преобразования, если происходит процесс абстрагирования, например такой, как детекция признака.
Для чего мозгу нужно так много замещающих схем, так много операций кодирования и перекодирования? Ведь любое преобразование грозит утратой точности. Почему же это свойство присуще всей нервной системе? Какую пользу дает перекодирование? Должно быть, ответ на этот вопрос состоит в том, что перекодирование повышает эффективность нервной системы.
Это перекодирование отнюдь не такая тривиальная операция, как та, с которой я столкнулся при использовании компьютера общего типа. Последним можно управлять только с помощью языка, который он понимает: пространственной или временной последовательностью «on» и «off», «да» и «нет», «вверх» и «вниз», системой имеющихся у него переключателей. Если имеется 12 таких переключателей, то мы должны запомнить размещение каждого из них для осуществления операции, которую мы хотим получить от компьютера. Следовательно, передача сообщения принимает вид серий
ABB AAA BAA BBB ААВ ABA AAB ABB ABA BBB ABA ABA и т. д.
Таким образом, стоящая перед оператором компьютера задача является в значительной мере задачей на запоминание длинного перечня позиций включения и выключения, рядов «двоичных» цифр.
011 000 100 111 001 010 001 011 010 111 010 010
Программисты должны были быстро овладеть этим сложным способом управления их инструментом: они подразделяли 12 переключателей на наборы по три и обозначали включение (В) целым числом, геометрически увеличивающимся справа налево…
 |
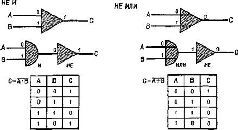 |
Рис. IV-1. Представление двоичных логических элементов: «и», «или», «не и», «не или». Входные сигналы 0 или 1 поступают в схему из двоичных элементов слева, где логически комбинируются для создания выходного сигнала справа. «Таблица соответствий» под каждой схемой перечисляет все возможные логические операции, которые могут еще быть выполнены. Элементы «не и» и «не или» образуются комбинациями элемента «не» с «и» и «или» (Evans, Computer Logic and Memory, 1966).
 |
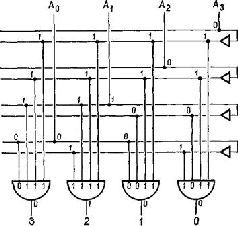 |
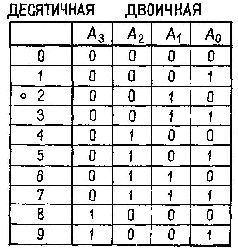
Рис. IV-2. Превращение двоичной системы в десятичную с помощью схемы, состоящей из 4 элементов «не» и «и». Таблица соответствий справа содержит двоичный эквивалент для десятичных цифр (от 0 до 9). Схема демонстрирует принцип декодирования двоичных чисел на примере декодирования числа 2. Сигнал на каждом из пронумерованных выходов будет равен 0, пока на все входы подается 1. В этом примере единица подается на вход А! третьего справа логического элемента «и», обозначенного цифрой 2. Таким образом двоичное число 0010 декодируется в число 2 десятичной системы (см. Evans, 1966).
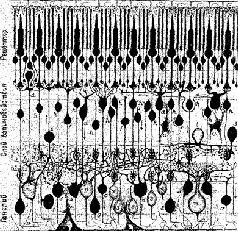 |
Рис. IV-З. Схема сетчатки приматов, показывающая типы нейронов и их синантические связи Метод Гольдзди. (Polyak, 1941).
Таким образом, в каждой триаде включение указывает на 4-2-1. Когда одновременно включаются два или три переключателя, то они представляют сумму целых чисел. Так,
0 означает AAA
1 означает ААВ
2 означает ABA
3 означает ABB
4 означает ВАА
5 означает ВАВ
6 означает ВВА
7 означает ВВВ
и любую последовательность из 12 включений и выключений можно описать и запомнить в виде четырех цифр, например представленные ранее последовательности превращаются в 3047, 1213, 2722. Это преобразование, названное «восьмиричным» кодированием двоичной системы, удивительно экономно. Каким же образом стало возможно столь замечательное достижение? (Этот -совершенный способ нелегко приобретается нервной системой; благодаря повторению процесса обеспечивается классификация по иерархическим схемам, а классификация – наиболее фундаментальная логическая процедура.) Чтобы получить ответ, сформулируем проблему более конкретно: каким образом может происходить преобразование одной структуры, сложность которой яредставлена
Таким образом, перекодирование оказывается чрезвычайно эффективной частью процесса памяти, для осуществления которого конструкция нервной системы, по-видимому, великолепно .приспособлена. Формы перекодирования, которые возможны в нервной системе, фактически безграничны. Тем не менее можно выделить несколько классов кодов.
Мы уже разделили коды на два класса: коды, предполагающие дискретные импульсы нервного разряда, и коды, предполагающие микроструктуру устойчивого состояния, создаваемую нервными соединениями.
Проблема импульсного кодирования не сводится к вопросу о присутствии или отсутствии сигнала; импульсный код может использовать множество различных параметров, которые характеризуют последовательности залпов.
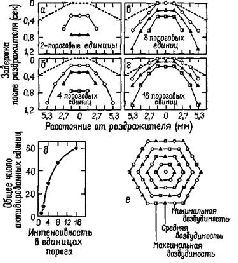 |
Рис. IV-4. Кодирование интенсивности и места раздражения, осуществляемое параллельными волокнами. Пространственное представительство показано в соответствии с тем, что было обнаружено в тактильных афферентах задней лапы кошки, когда последняя возбуждалась механическими залпами, приложенными к одной точке; диаграмма основывается на количественных экспериментальных данных. Если число равноотстоящих рецепторов с 3 уровнями возбудимости соответствует схеме е, то раздражитель интенсивностью в два порога вызывает некоторое число импульсов в стороне от раздражаемой точки и через какое-то время после раздражения а. Пунктирная линия обозначает латентный период, обусловленный временем распространения механической волны по лапе. Более сильные раздражители вызывают ответы, показанные на рис. б-г. Общее число импульсов в ответ на каждую интенсивность раздражителя представлено в д (Gray and Lai, 1965).
Д. Перкел и Т. Буллок (Perkel and Bullock, 1968), изучал сигналы, посылаемые электрическими органами некоторых рыб, установили следующие типы кодирования. При той или: иной комбинации условий изменяется длительность латентного периода, продолжительность спайковых взрывов, общая вероятность разрядов и вариация этой вероятности, увеличивается или уменьшается частота разрядов или скорость ее изменения. Такими условиями являются искажения электрического поля, создаваемые теми же самыми сигналами, посылаемыми электрическими органами, – искажения, регистрируемые системой боковой линии рыбы, этот процесс сходен с радарным.
Кроме этих организованных во времени кодов, имеет место и пространственное кодирование, когда множество параллельных линий – нервных волокон – продуцируют то, что Перкел и Буллой назвали обработкой «по ансамблю», которая зависит от различий в распределении последовательностей импульсов вдоль линий (см. рис. IV-4).
Пространственное кодирование и особенно кодирование, опирающееся на различия в синхронности разрядов у соседних групп нервов, тесно связано с целой группой процессов неимпульсного кодирования – микроструктурами медленных потенциалов, обеспечивающих устойчивое состояние, которые вносят существенный вклад в активность серого вещества мозга млекопитающих. Как говорят Перкел и Буллок, должно существовать какое-то считывающее устройство, какой-то механизм в центральной нервной системе, который считывает сообщения, закодированные в импульсной форме. В этих главах утверждается, что такое считывание возникает на местах соединений нейронов в процессе образования микроструктур медленных потенциалов, мгновенных состояний, возникающих в результате взаимодействия между совокупностями сигналов, действующих по соседству и кодируемых последовательностью импульсов.
Чтобы быть эффективными, большинство кодов нуждается в некотором стабильном уровне Работа Бернса и других, вне веяного сомнения, установила, что активность центральной нервной системы обладает такой стабильностью Нервная ткань спонтанно генерирует электрические потенциалы Мозг, как и сердце, непрерывно пульсирует И, так же как в сердце, такую пульсацию вызывают медленные потенциалы, а возникновение этих последних зависит от определенных констант химического окружения, в котором находится пульсирующая 1кань (рис IV-5).
 |
Рис IV-5. Церебральная симфония (Verzeano et al, 1970).
Серии тщательно проведенных исследований в лаборатории Бернса (1958) дали исчерпывающий ответ на вопрос, который долгое время оставался фантастическим: может ли мозг сохранять активность даже в том случае, если он полностью изолирован (нейронально) от другой нервной ткани? Результаты этих экспериментов, как это часто бывает, не подтвердили полностью ни представления о том, что активность головного мозга «спонтанна», ни представления о мозге как находящейся в покое tabula rasa, на которую записывается сенсорный опыт. Берне обнаружил, что даже у неанестезированного животного изолированная полоска коры остается неактивной до тех пор, пока к ней хотя бы на короткое время не будет приложено электрическое раздражение; другие же данные (Echlin et al., 1952; Gerard and Joung, 1937; Henry and Scoville, 1952; Ingvar, 1955; Libet and Gerard, 1939) указывают на то, что спонтанная активность существует и в таких препаратах. В любом случае, даже если принять осторожный вывод, сделанный Бёрнсом, несколько сильных электрических раздражителей, приложенных к поверхности коры, вызывают серию разрядов нейронной активности, которая обычно продолжается в течение многих минут (или даже часов) после прекращения стимуляции.
Периодические волны возбуждения могут быть получены и в диффузно организованной нервной ткани при ее электрическом раздражении. Они сходны с волнами возбуждения, возникающими в неанестезированной коре головного мозга в ответ на воздействие нескольких редких стимулов. Эффекты, длящиеся многие часы, наблюдались после короткой стимуляции интактной актинии (Batham and Pantin, 1950). Недавно был описан люминесцентный ответ у морских «анютиных глазок» (вид цветного коралла): после серий раздражений эти колонии начали люминесцировать спонтанно, а не только в ответ на стимуляцию. Для объяснения этого явления следует обратиться к механизму медленных изменений состояния нервной ткани (элементарной форме памяти, связанной с медленными потенциалами?) Эти изменения обусловлены влиянием окружающей среды и зависят, разумеется, от предшествующей активности организма. Но они также имеют свои внутренние закономерности и свой собственный ритм активности, который вызывает повторные изменения состояний нервной ткани, что делает их в каждый момент времени лишь частично зависимыми от влияний окружающей среды.
Короче говоря, принято считать, что группы нейронов того типа, которые найдены в коре головного мозга, в отсутствие непрерывного сенсорного воздействия находятся в состоянии покоя. Однако эти группы нейронов могут легко приходить в состояние- возбуждения и обнаруживать длительную активность. Значит, можно считать, что во время «покоя» они находятся в состоянии ниже порога непрерывного самовозбуждения. У интактного млекопитающего есть механизм, который поддерживает возбуждение центральной нервной системы выше этого уровня покоя. Таким механизмом является спонтанный разряд рецепторов.
Р. Гранит (1955) подробно рассказал о том, каким образом им «завладела мысль, что спонтанная активность является составной частью работы сенсорных систем». Он проследил историю этого вопроса от ранних наблюдений лорда Э. Эдриана и И. Зоттермана (1926), Э. Эдриана и Б. Мэттьюза (1927а, б), выполненных на мышцах и препаратах зрительного нерва, до его собственных разносторонних экспериментальных исследований. Более того, его данные подтверждают предположение, что эта «спонтанная» активность органов чувств делает их одним из наиболее важных «энергизаторов», или активизаторов, мозга. Сейчас мы можем добавить к этому, что, вероятно, эта спонтанная активность является той основой, тем уровнем, на котором и по отношению к которому осуществляется нейронное кодирование. Берне также представил данные, подтверждающие такое предположение (1968). С помощью микроэлектродов он обнаружил, что примерно 1/3 большого числа обследованных им клеток мозга в течение всего времени, пока он вел от них запись, показала устойчивость средней частоты своих разрядов. Эти нейроны реагировали на стимуляцию либо возрастанием частоты разрядов, либо их торможением. Всякий раз вслед за этим наступал период, во время которого активность нейрона менялась реципрокно реакции. В результате происходила компенсация изменений средней частоты разрядов нейрона, вызванных стимуляцией. Таким образом, эти клетки создают мощную стабильную базу, от которой зависит основная характеристика кодирования и перекодирования: пространственные структуры возбуждения могут возникать за счет возрастания спонтанной активности в одном месте и одновременного ее торможения в другом.
Способность нервной ткани генерировать спонтанную активность имеет более широкое значение, чем просто создание фона для более активных операций нервной системы. Имеются надежные данные, говорящие о том, что спонтанная активность нейронных групп может быть использована в работе того или иного кодирующего механизма. Например, можно показать, что при определенных условиях (при применении постоянного тока, вызывающего поляризацию нервной ткани) нейроны «запоминают» частоту раздражения, которое многократно применялось и вызывало у них реакцию вовлечения. Когда впоследствии частота раздражения менялась, нейроны воспроизводили частоту не нового, а многократно действовавшего первоначального раздражителя (Chow, 1964, Chow and Dewson, 1964; Dewson, Chow and Engel, 1964).
Впрочем, исследования Д. Пурпура (1962) пролили свет на гибкость этой способности к изменениям. В его экспериментах электрической стимуляции подвергалась какая-то группа клеток мозга, а запись проводилась от одиночных соседних нейронов.
Одновременно поляризирующий ток был приложен к нейрону, от которого производилась регистрация. Результаты эксперимента показали, что в условиях постоянной электрической стимуляции соседних нейронов ответ нейрона (6 разрядов в секунду) остается постоянным, несмотря на его поляризацию. Таким образом, используемая Пурпурой методика позволила ему показать, что распределение сигналов, подходящих к нейрону, от которого производилась регистрация, в каждый момент времени меняется таким образом, что компенсирует артефакт, вызываемый поляризацией. Это означает, что система нервных путей, через которые вызывается ответ нейрона, не фиксирована, она перестраивается в зависимости от условий (рис. IV-6).
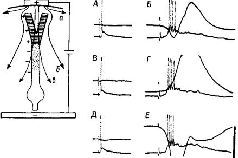 |
Рис. IV-6. Независимость изменений в вызванных кортикальных ответах и внутриклеточной активности нейрона пирамидного тракта, возникающих в ответ на поляризацию слабыми токами поверхности коры головного мозга (50 мкА/мм2). Верхний канал регистрирует поверхностные ответы на раздражение вентролатерального ядра таламуса. На конфигурации внутриклеточно записанной активности не влияют значительные изменения поверхностных вызванных ответов. А – антидромный спайк с хорошо выраженной задержанной деполяризацией; В – паттерны синаптического вовлечения во время стабилизированной фазы усиливающегося ответа; В и Г – во время слабой поверхностной анодной поляризации; Д и Е – во время поверхностной катодной поляризации. Калибровка: 50 мв, 20 мсек. Диаграмма вверху слева показывает вероятное распределение токов во время слабой анодной поляризации: а – фракция электрического потока; в – часть электрического потока, направленная вовнутрь окончаний апикальных дендритов и вовне, через проксимально расположенные дендритные зоны. Эффект этого тока не наблюдается на уровне сомы клетки при слабых интенсивностях, как показано в Г (Purpura, 1967).
Всякий раз, когда в каком-либо участке нервной системы спонтанная активность становится достаточно стабильной и при этом начинает управлять активностью других нейронных групп, она рассматривается в качестве пейсмекера. Некоторые пейсмекеры, такие, как пейсмекер, регулирующий сокращения сердечной мышцы, являются врожденными и функционируют на протяжении всей жизни индивида. Они определяют взаимоотношения организма с его окружением и тем самым он становится чем-то большим, нежели простым коммутатором, выполняющим требования своего жизненного опыта.
Другой тип пейсмекеров отличается несколько меньшим постоянством, такие пейсмекеры формируются в виде «временных доминантных очагов» в результате конвергенции на них возбуждения. Простой опыт, в котором такой временный доминантный очаг возбуждения был получен экспериментально, был проведен много лет назад Залманзоном (см. Ухтомский, 1926). У собаки был выработан условный рефлекс: в ответ на определенный тон она поднимала правую заднюю лапу. После выработки условной реакции была произведена операция, в результате которой правая двигательная зона коры, контролирующая левую сторону тела, была обнажена для последующего химического воздействия. Затем, во время осуществления условной реакции, на область, которая контролирует левую переднюю лапу, накладывался кусочек фильтровальной бумаги, смоченной стрихнином. Это сразу привело к тому, что собака меняла «отвечающую» конечность. Теперь на условный сигнал она поднимала не правую заднюю, а левую переднюю лапу. Это изменение реакции обычно объяснялось тем, что возникший в процессе выработки условной реакции очаг, доминировавший над другими функциями моторной коры, теперь оказался подавлен, благодаря новому «временному доминантному очагу», возникшему в этой области мозга в результате химического раздражения стрихнином (см. рис. IV-7, суммарная схема методов, используемых для получения доминантных очагов).
 |
Р и с. IV-7. Методы выработки условных связей, которые были использованы различными исследователями для формирования и изменений доминантных очагов в головном мозгу. В тексте пример относится к эксперименту Залманзона (John, 1967).
Но большинство пейсмекеров характеризуется цикличностью. В своей простейшей форме они выполняют функцию биологических часов (Richter, 1955; Pittendrigh, 1960). В системах, у которых выход взаимодействует с возбуждением, непрерывно поступающим на вход, пьйсмекеры выполняют функцию нейронных заслонов, которые лишь периодически пропускают несколько импульсов (как в кинокамере, Lindsley, 1961), и сканирующих устройств, которые систематически исследуют пространственное распределение входного сигнала, как в телевизоре (McCulIoch, 1965). Когда выход системы регулируется менее периодически и он при этом запрограммирован, то она описывается как система с «произвольными характеристиками».
В течение многих лет было невозможно провести различие между потенциалами и теми пейсмекерами, которые связаны с явлением нарастания и убывания нейроэлектрических состояний, возникающих в микроструктуре соединений. Но, как уже было отмечено, Соколов (1970) показал, что в определенных лабораторных условиях механизм генерации аксонных нервных импульсов может действовать относительно независимо от изменений медленных потенциалов мембраны тела клетки. Согласно этим данным, генераторный потенциал возникает внутри структуры тела нейрона, а не на его поверхности и, следовательно, он не может быть отнесен непосредственно за счет изменений характеристик мембранного потенциала нервной клетки и дендритных влияний на них (см. также рис. IV-8).
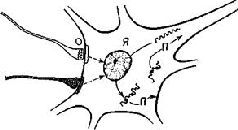 |
Рис. IV-8. Модель внутриклеточного контроля нейронных ритмов. Я – ядро; П – полипептид или протеин. Волнистая линия изображает переносчик (информационный) РНК, которая достигла или не достигла рибосом (). Соединение между субсинаптической мембраной, ядром и мембраной тела клетки, или аксоплазмой, показано сплошными стрелками. Синаптические пузырьки (О) показаны на нервном окончании вверху слева (см. Strumwasser, 1967).
Другая интересная диссоциация между генерацией импульсной активности и флуктуациями электрического состояния наблюдается во время одной из фаз сна. Эта фаза характеризуется быстрыми движениями глаз, часто подергивающимися движениями тела (ото можно увидеть, если понаблюдать за спящей собакой) и электрическими ритмами, которые могут быть записаны от мозга и которые неожиданно сходны с ЭЭГ бодрствующего состояния. Будучи разбужен на этой стадии сна, человек почти всегда сообщает, что он видел сны, тогда как у людей, проснувшихся во время других фаз сна, такие сообщения бывают редки.
Во время бодрствования у большей части нейронов мозга кошки (Evarts, 1967) повторяющаяся регулярность в разрядах не наблюдается. Во время обычного сна эти же клетки разряжаются взрывами спайков с периодами относительной неактивности между ними. Эти интервалы примерно соответствуют флуктуациям, одновременно записываемым макроэлектродами. Однако во время парадоксальной фазы сна нейроны дают интенсивные разряды с интервалами, повторяющимися примерно четыре раза в секунду, но электрические записи с макроэлектродов (ЭЭГ) не отражают этот доминирующий ритм. Это объясняется тем, что, хотя нейроны, разряжаясь, группируют свои спайки в четко выраженную временную структуру, они по фазе разряжаются независимо друг от друга, их активность не коррелирует с активностью соседних нейронов. Следовательно, корреляция спайковой активности отдельных нейронов не зависит от самого механизма генерации нервных импульсов. Поэтому должен существовать специальный механизм, осуществляющий связь спайковых разрядов соседних нейронов. Скорее всего, функцию этого механизма эффективно выполняет микроструктура медленных потенциалов, возникающих на контактах между нейронами. Таким образом, маловероятно, что устойчивое состояние и дискретные нейронные процессы являются взаимосвязанными процессами и отражают друг друга. Правила преобразования, которые позволяют перекодировать один процесс в другой, должны быть определены эмпирически для каждого из тех условий, которые интересуют исследователей. На данном этапе исследования любые общие утверждения относительно типов таких преобразований являются, вероятно, преждевременными.
Такая потребность в эмпирической информации, сообщающей о конкретных примерах преобразования дискретного процесса в непрерывный и наоборот, не исключает возможности создания гипотез о типах преобразований, которые, вероятнее всего, характезируют обычные операции перекодирования. Такие гипотезы необходимы еще и потому, что они определяют направление дальнейших исследований. По определению они будут соответствовать только категории обратимых преобразований, поскольку необратимые операции условны и, следовательно, должны быть просто открыты в эксперименте. Практически любой психологический процесс, который может быть описан словами, мог бы быть нейронно закодирован посредством необратимой операции. Однако остается открытым вопрос, действительно ли это является механизмом, пригодным для объяснения любого конкретного случая.
Как станет очевидным из последующих глав, обратимые преобразования, и в первую очередь некоторые классы линейных преобразований, особенно полезны для объяснения психологических явлений. Ряд возможных правил обратимого преобразования был легко установлен, и сейчас эти правила настолько проверены, что, руководствуясь ими, компьютеры могут помогать нам анализировать матрицы с включенными в них данными. Вот несколько примеров:
1. Суммация (или вычитание) следующих друг за другом нервных импульсов преобразуется в амплитудные флуктуации состояния постсинаптической микроструктуры.
2. Последовательно возникающие состояния медленных потенциалов в -микроструктуре соединений достаточно хорошо коррелируют с потенциалами, управляющими генераторным потенциалом нейрона.
3. Нервные импульсы, одновременно достигающие соседних участков, пространственно суммируются, то есть имеют место взаимодействия по-соседству в виде сложения или вычитания.
4. Когда два источника одновременно изменяют состояние микроструктуры медленного потенциала, между ними возникает корреляция, которая вновь может декодироваться в нервные импульсы.
Имеющиеся данные позволяют предположить, что ряд нейронных механизмов, многие из которых существуют с рождения, подчиняется этим правилам. Поэтому в следующей главе будут подробно рассмотрено, какие формы принимают некоторые из основных модулей нервной системы.
Если каждая группа нейронов осуществляет операцию перекодирования получаемых на входе воздействий, то должны существовать изменчивые динамические структуры организации нейронов. Перекодирование – удивительно мощный адаптивный и конструктивный инструмент, которым пользуется организм, чтобы действовать во внешнем мире и воздействовать на него. Некоторые операции кодирования сохраняют функциональный изоморфизм и обратимы при условии, что выполняются соответствующие преобразования. Такие преобразования возникают в результате тормозных взаимодействий в местах соединения нейронов.
| © 2024 Библиотека RealLib.org (support [a t] reallib.org) |